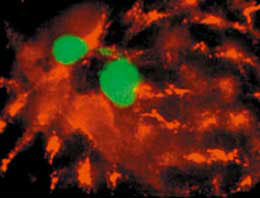
Merosomen
von Petra Stöcker (Laborjournal-Ausgabe 12, 2007)
Durstige Weibchen stellen im Allgemeinen keine ernsthafte Bedrohung dar, schon gar nicht für den Wirt. Anders verhält es sich im Falle der weiblichen
Anopheles-Mücke. Beherbergt sie blinde Passagiere der Gattung
Plasmodium, kann ihr Stich eine lebensbedrohliche Malaria-Infektion im Säugetier-Wirt auslösen.
Gefährlich für den Menschen sind nur wenige
Plasmodium-Arten:
Plasmodium vivax beziehungsweise
ovale verursacht die so genannte Malaria tertiana,
Plasmodium malariae die Malaria quartana. Die weiteste Verbreitung und zugleich höchste Letalität findet sich bei der Malaria tropicana, hervorgerufen durch eine Infektion mit
Plasmodium falciparum. Für diese Erreger sind Stechmücke und Mensch die einzigen Wirte. Der komplizierte Lebenszyklus der einzelligen Parasiten ist bis auf wenige Lücken gut erforscht. Im Lauf ihres Lebens nehmen sie unterschiedlichste Gestalten an.
Geschickte Tarnung
Mit dem Speichel infizierter Mücken werden die Plasmodien in Form so genannter Sporozoiten in den menschlichen Blutstrom geschwemmt. Innerhalb weniger Minuten erreichen sie die Leber, in deren Parenchymzellen sie ihren ersten vegetativen Vermehrungszyklus durchlaufen. Während dieses "prä- (exo-) erythrozytären Zyklus", der je nach Plasmodien-Spezies zwischen zwölf und fünfzig Tage dauert, entwickeln sich die Parasiten zu Leberschizonten, die wiederum in tausende von Merozoiten zerfallen.
Die frei beweglichen Merozoiten befallen schnell Erythrozyten. In deren Inneren wachsen sie und teilen sich. In jedem Erythrozyten entsteht über die Ringform (Trophozoit) ein neuer Schizont, der sich wiederum in viele Merozoiten teilt, die in den Blutstrom freigesetzt werden und andere rote Blutkörperchen befallen.
Damit beginnen die Malaria-Symptome, denn der periodisch wiederkehrende Zerfall der Erythrozyten geht mit Fieberschüben einher. Um zu den Erythrozyten zu gelangen müssen die hepatischen Merozoiten das Lumen der Leber-Blutgefäße (Sinusoide) erreichen. Dabei passieren sie das gefensterte Sinusoid-Endothel und den "Disse-Raum" (Raum zwischen dem durchbrochenen Endothel der Lebersinusoide und den Leberzellplatten, der den unmittelbaren Kontakt zwischen Blut und Leberzellen ermöglicht).
Bislang war unklar, wie die Merozoiten dabei der Phagozytose durch in den Leber-Sinusoiden patroullierende Makrophagen, den Kupffer-Zellen, entwischen können. Eine Antwort bietet das Hamburger Forscherteam um Volker Heussler vom Bernhard-Nocht-Institut (BNI) für Tropenmedizin (Science 313:1287).
Sie verfolgten die Entwicklung von
Plasmodium berghei, einer Nagetier-infizierenden Spezies, an einer in vitro-Kultur von HepG2-Zellen. Der zwei bis drei Tage dauernde Entwicklungszyklus dieser Plasmodien findet in einer parasitären Vakuole statt, in der sich der Sporozoit zuerst in einen großen, vielkernigen Schizonten und dann in tausende von Merozoiten differenziert.
Nach insgesamt 63 bis 70 hpi (
hours post infection) hatte sich die Anzahl Parasiten-gefüllter adhärenter Zellen um sechzig Prozent verringert, doch schwammen nur wenige freie Merozoiten im Zellkultur-Überstand, wo man sie nach dem vermeintlichen Zerplatzen der Leberzellen erwartet hätte. Stattdessen schwammen den Hamburgern zahlreiche dicht mit funktionsfähigen Merozoiten bemannte, unversehrte Leberzellen vor die Linse des Mikroskops.
Merozoiten-Fähren
Unerwarteterweise fanden sie Merozoiten auch in runden Vesikeln ohne Zellkern, die von außen an die Zellmembran von Leberzellen gebunden waren. Die Leberzelle pumpt die Parasiten in Membransäckchen (Merosomen), die später von der Zelle abgeschnürt und in ein benachbartes Blutgefäß entlassen werden.
Die so umhüllten Parasiten werden von den zahlreichen Fresszellen der Leber nicht als Eindringlinge erkannt. Im Merosom – ihrem trojanischen Pferd – gelangen die Parasiten aus dem Lebergewebe sicher zurück in die Blutbahn. Einmal in den Blutkreislauf entlassen, zerstören die Parasiten das Merosom, werden frei gesetzt und beginnen mit der Erythrozyten-Infektion. Der Übergang in diese "Blutphase" markiert den Beginn der Malaria-Erkrankung.
Die Merosomen-bildenden Wirtszellen zeigten in vitro die klassischen Merkmale des apoptotischen Zelltodes, wie Kondensation des Zellkerns, Verlust des mitochondrialen Membranpotentials und Cytochrom c-Freisetzung. Allerdings wird hier der Zelluntergang nicht durch Caspasen, sondern durch andere Cystin-Proteasen eingeleitet. Auch wird die DNA in den kondensierten Wirtszellkernen nicht fragmentiert.
Schutz vor Phagozytose
In der Membran von abgelösten Wirtszellen und Merosomen blieb die asymmetrische Verteilung von Phosphatidylserin (PS) erhalten – Kennzeichen lebender Zellen. Während der Apoptose wird Phosphatidylserin in die äußere Membranschicht verlagert. Dies ist Teil des Prozesses, welcher die Zelle für die Phagozytose markiert. Phagozyten orientieren sich an dem von sterbenden Zellen auf der Oberfläche exprimierten Phosphatidylserin, um "tot" von "lebend" zu unterscheiden.
Hepatische Merozoiten unterbinden die Phosphatidylserin-Expression in ihren "
Shuttle-Zellen", indem sie die dafür notwendige Ca
2+-Freisetzung aus Mitochondrien in das Zytoplasma verhindern und die intrazelluläre Ca
2+-Konzentration niedrig halten. Damit bleiben die Zellen und ihre Fracht vor Phagozyten-Fraß verschont.
Mit dieser geschickten Tarnung gelangen die Parasiten unentdeckt zu den Erythrozyten im Blutstrom. Die detektivische Arbeit der Hamburger Wissenschaftler zeigt neue Möglichkeiten auf, die Invasion von Erythrozyten und damit den Beginn der Malaria-Erkrankung zu verhindern.
Letzte Änderungen: 08.02.2008