Endozytose-Adapterproteine
von Mario Rembold (Laborjournal-Ausgabe 04, 2014)
Die Zellmembran besteht bekanntlich aus weit mehr als einer schnöden Doppelschicht aus Lipiden, die lediglich das Innere vom Äußeren trennt. Vielmehr ist sie gespickt mit Kanälen, Pumpen und Rezeptoren, die gezielt Moleküle in die Zelle hinein und aus ihr heraus befördern – oder wenigstens Informationen durch die Membran weitergeben.
Ein Weg in die Zelle führt über die Endozytose: Ein Teil der Membran stülpt sich nach innen und schnürt sich ab; im Zytoplasma schwimmt nun ein Vesikel, das die außen aufgenommenen Stoffe enthält. Auf diese Weise können Nährstoffe ins Zellinnere gelangen, oder holen sich Neuronen ausgeschüttete Transmitter zwecks Recycling aus dem synaptischen Spalt zurück – um nur zwei Beispiele zu nennen. Doch auch manch ein unerwünschter Gast verschafft sich mittels Endozytose Zugang zur Zelle. Zahlreiche Viren missbrauchen den Mechanismus, um an die Membran anzudocken und den Wirt zu entern.
Dellenbildung
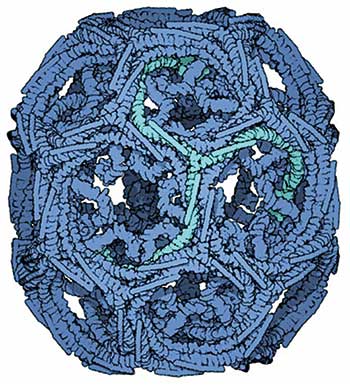
Es gibt unterschiedliche Mechanismen der Endozytose, wobei die Vesikelbildung unter Clathrin-Beteiligung am besten verstanden ist. Ein einzelnes Clathrin ist ein Hexamer aus drei schweren und drei leichten Ketten und nimmt die Form dreier radiär angeordneter Kreisbögen an. Lagern sich mehrere dieser Proteine an der Zell-inneren Seite der Membran aneinander, bilden sie ein korbartiges Geflecht, das sich nach innen krümmt und die Doppellipid-Schicht dabei mitzieht. Ein von Calthrin ausgekleideter (clathrin coated) Membranbereich formt also eine konkave Delle und stülpt sich immer stärker nach innen, je mehr Clathrin sich anlagert. Aus den Dellen (coated pits) werden schließlich sphärische Gebilde, denn durch die Krümmung des Clathrin-Geflechts entsteht am Ende eine Kugel, die sich von der Membran nach innen hin abschnürt. Fertig ist das Clathrin-coated Vesikel.
Ganz so einfach wie hier dargestellt, ist die Clathrin-abhängige Endozytose aber nicht. Denn Clathrin lässt sich nicht einfach so an der Membran befestigen. Hierfür braucht es Adapterproteine, die den Kontakt vermitteln. Prominentestes Beispiel ist der heterotetrameric adaptor protein complex AP2. AP2 besteht aus vier verschiedenen Untereinheiten und bindet an PIP2, einem Phospholipid, das fast ausschließlich in der inneren Schicht der Zellmembran vorkommt. Damit ist AP2 fest auf der zytosolischen Seite der Membran verankert. Aus der zentralen AP2-Domäne ragen zwei Schwänzchen mit Bindungstellen für Clathrin ins Zytosol hinein. Anscheinend genügen zwei nebeneinander liegende AP2-Komplexe mit einem gebundenen Clathrin, um nachfolgend weitere Clathrin-Proteine und AP2-Adaptoren anzulagern und damit Endozytose zu initiieren.
AP2 verfügt außerdem über Domänen, die an die zytoplasmatische Seite bestimmter Transmembranproteine binden können: die Cargo-Rezeptoren. An diese Rezeptoren wiederum dockt von zelläußerer Seite die Fracht an – also die Moleküle, die per Endozytose in die Zelle wollen. Auf diese Weise baut AP2 nicht nur eine Brücke zwischen Clathrin und Zellmembran, sondern stellt auch sicher, dass die aufzunehmende Fracht dort bleibt, wo die Endozytose stattfindet.
Mäuse können nicht ohne, Hefe wohl
Egal ob Tier, Pflanze oder Hefe – immer wieder stößt man auf Clathrin-abhängige Endozytose. Weniger konserviert ist hingegen die Rolle der Adapterproteine. AP2 scheint eine Schlüsselfunktion zu erfüllen, sollte man meinen – schließlich können AP2-defekte Mäuse keine Clathrin-Vesikel mehr bilden. Allerdings zeigt sich Hefe recht unbeeindruckt, wenn man die Bildung des AP2-Komplexes stört.
Tatsächlich können nämlich eine ganze Reihe von Proteinen Clathrin mit der Zellmembran und den Cargo-Rezeptoren verbinden. Obgleich in Säugerzellen der AP2-Komplex unverzichtbar ist, gibt es auch hier weitere Adapterproteine wie Intersectin 1 und 2, Epsin 1 bis 3 oder Dab2. Und Homologe zu einigen dieser Proteine finden sich eben auch in Saccharomyces, welche notfalls ohne funktionsfähigen AP2-Komplex die Bildung von Clathrin-Vesikeln unterstützen (für einen ausführlichen Überblick über die Rolle diverser Adapterproteine und die Initiation Clathrin-vermittelter Endozytose siehe J. Cell Biol. 203(5):717-25).
Pflanzen sind besonders
Andere Adapterproteine wiederum sind keineswegs derart ubiquitär in der Eukaryoten-Welt anzutreffen. So erschien im Februar eine Arbeit der Erstautorinnen Astrid Gadeyne (Ghent, Belgien) und Clara Sánchez-Rodríguez (Potsdam-Golm), die zusammen mit weiteren europäischen Kollegen einen Proteinkomplex in Arabidopsis charakterisierten, der ausschließlich in Pflanzen Clathrin-Vesikel bilden hilft (Cell 156(4):691-704).
Bereits zuvor hatten Mitglieder des Teams gezeigt, dass ein Protein namens TPLATE mit Clathrin interagiert. In der aktuellen Arbeit isolierten die Autoren TPLATE mittels Tandem-Affinitätsreinigung, um zu sehen, welche Proteine daran gebunden sind. Sieben an der Zahl identifizierten sie schließlich, die zusammen mit TPLATE einen Komplex bilden – den TPC (für TPLATE complex). Manipuliert man Proteine des TPC und beeinträchtigt damit die Funktionalität des Komplexes, so ist die Aufnahme bestimmter Cargo-Proteine in Arabidopsis gestört. Zwar ähneln zahlreiche Domänen des TPC solchen, die in Hefe und im Tierreich bei Adapterproteinen vorkommen, klare Homologe der beteiligten Proteine findet man jenseits der Pflanzenwelt aber nicht. Die Autoren schließen daraus, dass die Adapterproteine sich im Laufe der Evolution sehr stark auseinanderentwickelt haben müssen.
Letzte Änderungen: 07.04.2014