Kunstvolle Genmosaike
Spezial: Duale ifg-Mosaike
Andrea Pitzschke
Die Herstellung induzierbarer genetischer Mosaike, mit denen sich Genfunktionen in einzelnen Zellen untersuchen lassen, war in Wirbeltieren bisher sehr kompliziert. Mit einer neuen Technik geht es erheblich einfacher.
Wir alle sind genetische Mosaike. Obwohl wir aus einer einzigen befruchteten Eizelle hervorgingen, enthalten unsere Zellen keine völlig identischen Genome. Schon während der Embryonalentwicklung kommt es zur ein oder anderen Mutation. Die kann neutral, von Vorteil oder nachteilig sein. Je früher sie auftritt, desto mehr Tochterzellen sind betroffen.
Genetisch bedingte Krankheiten wie Krebs müssen daher nicht zwangsläufig erblich sein. Weitergereicht werden sie nur, wenn Keimbahn-Mosaike, also nicht-somatische Mosaike vorliegen. Elefantenmenschen (Proteus-Syndrom) liefern ein gutes Beispiel für somatische Mosaike. Sie „verdanken“ ihren Körperbau Mosaik-Mutationen im AKT1-Onkogen. Je nach Verteilung und Intensität wuchern manche Körperregionen massiv, einzelne Gliedmaßen bleiben hingegen völlig symptomfrei.
Tanzen einzelne Zellen im Gewebe durch eine Mutation aus der Reihe, können sie ihre nicht-mutierten Nachbarzellen beeinflussen, zum Beispiel durch Sekrete. Anders herum können aber auch die Nachbarzellen dafür sorgen, dass mutierte Zellen sich unauffällig benehmen und ihrer unkontrollierten Vermehrung Grenzen setzen.
Kompliziertes Wechselspiel
Dieses Wechselspiel zu durchschauen, ist für Forscher, die die Entwicklung von Organen oder auch Krankheiten untersuchen, nicht einfach. Noch werden die meisten Vergleichsstudien an vermeintlich genetisch homogenen Systemen durchgeführt, zum Beispiel normalen und mutierten Mäusen oder normalen und transgenen Zellkulturen. Diese geben jedoch keinen Einblick in potenzielle Einflüsse durch genetisch andersartige Nachbarzellen.
Genetische Mosaike ermöglichen den Vergleich von Organismen mit prinzipiell identischem genetischem Hintergrund. Die Organismen unterscheiden sich lediglich in der einen oder anderen Zelle, die mutiert ist oder ein Wunschgen exprimiert. Irreführungen durch sekundäre und nicht-zellautonome Effekte entfallen daher.
Möchte man wissen, welche Konsequenzen bestimmte Mutationen in Zell-Subpopulationen für ein ansonsten normales (Wildtyp-) Gewebe haben, muss man einzelne Mosaiksteinchen manipulieren und das veränderte Gesamtbild funktionell untersuchen. Anders als der klassische Mosaikleger, der zuerst unterschiedliche Steinchen sammelt und dann zusammenlegt, geht der Genetiker von einem „farblosen“ Grundgebilde aus, etwa einer Zellkultur oder einem Gewebe. Durch gezielten DNA-Umbau ändert er Form und Farbe einzelner Steinchen, respektive Zellen.
Sein Hauptwerkzeug ist hierbei die Cre-vermittelte Rekombination. Diese beruht auf der ursprünglich aus Bakteriophagen stammenden Rekombinase Cre und zwei kurzen DNA-Erkennungsmotifen (LoxP). Der Abstand der beiden Elemente voneinander ist relativ flexibel. Je ein Lox-Motiv upstream (links) und downstream (rechts) des gewählten DNA-Abschnitts bewirken, dass beim Eintreffen von Cre genau dieser Abschnitt hinausgeworfen und die entstandene Wunde verschlossen wird. Folgt auf das rechte Lox-Motiv ein Zielgen, gibt die Cre-vermittelte Deletion den Startschuss für dessen Expression. So kann etwa aus einer fluoreszierenden Zelle eine Onkogen-produzierende Zelle werden.
Gefloxte Mäuse
Solange die Rekombinase ruht, passiert nichts. Erst durch Kreuzung einer „gefloxten“ Zelllinie (zu deletierender DNA-Abschnitt flankiert von zwei Lox-Motiven) mit einer Cre-exprimierenden Zelllinie gelangt Cre zum Wirkort. Platziert der Genetiker vor das Cre-Gen einen gewebespezifischen oder induzierbaren Promoter, kann er die Rekombination räumlich und zeitlich kontrollieren.
In Organismen wie Drosophila, in denen Rekombinationen leicht stattfinden, sind Mosaik-Techniken zur Genfunktionsanalyse gut etabliert. Für humanmedizinisch relevante Aussagen muss aber zumeist die Maus als Modell herhalten. Hier ist es etwas schwieriger, genetische Mosaike herzustellen. Die für die Manipulation nötigen DNA-Konstrukte sind sehr umfangreich und entsprechend schwer im Genom unterzubringen.
Eine der gängigen Techniken ist die MADM-Strategie (Mosaic Analysis with Double Markers). Mit ihr gelang es 2005 einem amerikanischen Forscherteam erstmals, Mosaike in Mäusegeweben durch interchromosomale Rekombination zu erzeugen (Cell, 6;121(3): 479-92). Das Verfahren funktioniert allerdings nur mit konstitutiv aktiven Cre-Linien. Weil interchromosomale Rekombinationen seltene Ereignisse sind, tragen bei MADM nur sehr wenigen Zellen eines Gewebes die gewünschte genetische Manipulation.
Könnte man nicht einfach die Frequenz der Rekombinationsereignisse erhöhen, um genetische Mosaike zu erzeugen? Mit der bislang üblichen Methode für Genfunktionsanalysen, die auf der Mosaikbildung durch intrachromosomale Rekombination basiert, ist dies möglich. Mit einem gewebespezifischen Promotor und dem CreERT2-Liganden Tamoxifen wird die Rekombination am gewünschten Ort zu einer festgelegten Zeit ausgelöst.
Die anfängliche Euphorie wurde jedoch rasch gebremst, als sich herausstellte, dass die Rekombination des Reporter-Allels (zum Beispiel der Verlust der Fluoreszenz durch Deletion) und die Aktivierung des Zielgens nicht unbedingt korrelierten (Genesis, 51(6): 436-42). Welche Zellen des genetischen Mosaiks das Zielgen tatsächlich exprimieren und welche abnorme Zellform sie selbst und etwaige beeinflusste Wildtyp-Nachbarzellen annehmen, blieb mangels Signalstärke beziehungsweise Sensitivität weitgehend im Dunkeln.
Schwarz-weiß statt bunt
Die rekombinierten Mosaiksteinchen offenbarten ihre Identität erst in Immunostaining-Analysen. Dummerweise ist die dafür nötige Zellfixierung mit Lebendzell-Untersuchungen partout nicht vereinbar. Mehrere Reporter- und Zielgene für einen komplizierteren Mosaikbau zu kombinieren erwies sich weitgehend als Illusion. Sprich, die Mosaike waren nicht bunt, sondern nur schwarz-weiß – eigentlich nur grau-weiß.
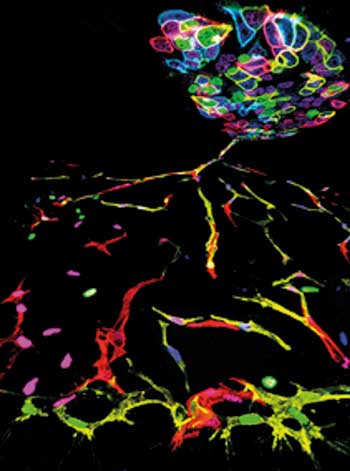
Die Gruppe des portugiesischen Angiogenese-Spezialisten Rui Benedito vom Spanish National Center for Cariovascular Research in Madrid stellte jetzt eine neue Strategie zur Herstellung genetischer Mosaike vor, die diese Hürden aus dem Weg räumt. Das Team benötigt nur zwei DNA-Konstrukte (iMb-Mosaik und iChr-Mosaik), um komplexe genetische Mosaike aus bis zu fünfzehn verschiedenen „Steinchen“, respektive Zellklonen zu erzeugen (Cell 170, 800-14).
Beide Konstrukte tragen die drei Lox-Motiv-Paare Lox1, Lox2 und Lox3. Auf der linken Seite sind Lox1, 2 und 3 nach einem starken Promotor platziert. Ihre jeweiligen Lox-Partner auf der rechten Seite sitzen vor dem ersten, zweiten sowie dritten Reportergen. Als Reportergene dienen im iMb-Mosaik-Konstrukt YFP, Tomato und Kate2, im iChr-Mosaik mCherry, eGFP sowie Cerulean. Die Reporter des iMb-Mosaiks sind mit dem Membran-Tag Mb fusioniert, die Reporter des iChr-Mosaiks mit dem Histon-Tag H2B; das Membran-bindende Peptid Mb führt die Reporter zur Membran, das Chromatin-bindende Peptid H2B in den Zellkern. An die Reporter können zusätzliche Gene angehängt werden, die untersucht werden sollen. Ein sich anschließendes Stoppsignal sorgt für das Ende der Expression.
Clevere Rekombination
Der Clou ist, dass die Cre-vermittelte Rekombination bei diesen Konstrukten zur zufälligen und sich gegenseitig ausschließenden Aktivierung je eines der drei Reportergene führt. Zielt die Rekombinase zum Beispiel auf das Lox1-Paar, versperrt sie unwiderruflich den Weg für mögliche Rekombinationen der Lox2- und Lox3-Paare. In diesem Fall wird also ausschließlich das erste Reportergen exprimiert.
Die Spanier platzierten die Mosaik-Kassetten in den Rosa26-Lokus von Mäusen, der häufig für die konstitutive, ubiquitäre Genexpression eingesetzt wird. Hieraus entstanden die Mäuse-Zelllinien iChr-Mosaik und iMb-Mosaik. Durch die Transfektion mit einem Cre-exprimierenden Plasmid ist mit diesen die Expression von drei Chromatin-lokalisierten (iChr-Mosaik) sowie drei Membran-lokalisierten (iMb-Mosaik) Fluoreszenzproteinen möglich.
In der F0-Generation folgen hieraus je drei verschiedenfarbige Markierungen an Zellkern sowie Zellmembran. Das Kern-Label erlaubt die Zellzählung, die Membranmarkierung gibt Auskunft über die Zellform. Kreuzt man Zelllinien, die eines der beiden Konstrukte tragen, ergeben sich insgesamt fünfzehn Kombinationsmöglichkeiten für die F1-Generation: Sechs resultieren aus den jeweils drei Reportergenen von Vater und Mutter. Die weiteren neun Kombinationen treten in Zellen auf, die Reportergene beider Konstrukte koexprimieren. Dies könnte zum Beispiel aus der Rekombination des Lox1-Motivs auf dem ersten, sowie der Rekombination des Lox3-Motivs auf dem zweiten Konstrukt resultieren. Als Konsequenz würden Zellen kernlokalisierte YFP-Signale und membranständige Cerulean-Signale aussenden.
Diese sogenannten Dual Induced Fluorescent Functional Gene Mosaics (ifg) eignen sich für epistatische Analysen. Die zu testenden Kandidatengene werden hierzu mit einem der Reportergene fusioniert. Wer in den Mosaiken die Häufigkeit der fünfzehn Möglichkeiten nicht dem Zufall überlassen will, kann die Lox-Paare entsprechend platzieren: Je näher die beiden Partner eines Lox-Paares beieinander liegen, desto wahrscheinlicher kommt es zur Rekombination. Das heißt, bei drei einzubauenden Zielgenkassetten kommt die kürzeste an den Anfang, die mittlere ins Zentrum, die längste ans Ende.
Bunte Mosaikmuster
Die Gruppe testete die Strategie unter anderem in embryonalen Stammzellen von Mäusen mit normaler oder verstärkter Notch- beziehungsweise VEGFR2-Signalleitung. Die Forscher beobachteten nach der Transfektion mit dem Cre-Plasmid bunte Mosaikmuster bei der Lebendzell-Analyse unter dem Mikroskop. Zusätzlich platzierten sie das Notch-Gen hinter eines der Reportergene und generierten mit diesem Konstrukt transgene Mäuse. Die iChr-Notch-Mosaik-Mauslinie kreuzten sie schließlich mit mPolr2a-CreERT2 Mäusen, um die Rekombination durch die gewebespezifische, Tamoxifen-induzierbare Expression von Cre in CreERT2-Mäuse zu steuern.
Hierzu erhielten Embryonen der gekreuzten Mäuse eine Tamoxifen-Spritze. Vier Tage später analysierten die Spanier die Proliferation und Differenzierung der Zellen während der Neuro- und Angiogenese – zwei zelluläre Prozesse die von den Notch und VEGFR-Signalwegen abhängen. Ein Ergebnis dieser Analyse war, dass einzelne Nervenvorläuferzellen offensichtlich recht schnell proliferieren, sich aber nur langsam weiter differenzieren – wodurch die Neurogenese länger aufrecht erhalten wird.
Theoretisch müsste sich das ausgetüftelte System der spanischen Gruppe erweitern lassen, um die Zahl der Kombinationsmöglichkeiten weiter zu erhöhen. Hierfür bräuchte es nur ein weiteres Lokalisationssignal, etwa für Mitochondrien, sowie drei zusätzliche Reportergene.
Letzte Änderungen: 03.11.2017