Von der Klonierung zur Assemblierung
Werkzeuge der synthetischen Biologie
Frederique Wieters
Eine wesentlicher Prozess in der synthetischen Biologie ist der Zusammenbau von DNA-Fragmenten zu größeren Einheiten, die eine gewünschte Funktion erfüllen. Verschiedene Assemblierungs-Techniken und Standards sollen den Forschern bei dieser kniffligen Aufgabe helfen.
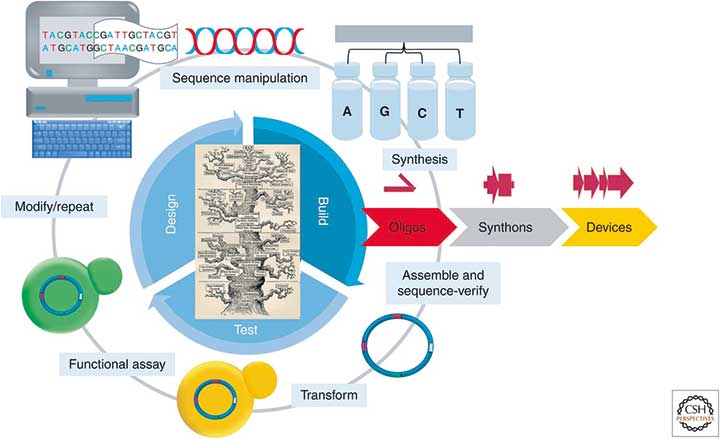
Das 21. Jahrhundert könnte nach Meinung vieler Experten das Jahrhundert der Biotechnologie werden. Hauptverantwortlich hierfür sind moderne DNA-Sequenzier- und Synthese-Methoden, die völlig neue Wege für die Untersuchung und Manipulation von Organismen eröffnen. Ein Beispiel ist die synthetische Biologie, die Molekularbiologie, Ingenieurswissenschaften, Informatik, Nanobiotechnologie und organische Chemie mit dem Ziel vereint, neuartige biologische Systeme herzustellen. Ausgangspunkt hierfür sind synthetische DNA-Nukleotide, die zu größeren DNA-Bausteinen (Synthons), Genen oder kompletten Genomen zusammengesetzt (assembliert) werden und künstlichen Zellen oder Organismen neue Eigenschaften verleihen sollen.
Wissenschaftler der synthetischen Biologie verfolgen zwei unterschiedliche Strategien. Eine ist die Integration synthetischer Systeme in biologische Systeme. Die andere zielt auf die Verkleinerung des Genoms ab, um die Systemkomponenten auf das absolute Minimum zu reduzieren. Diese minimalen Organismen werden als Gerüst für sogenannte Bioparts genutzt, die biologische Schaltkreise erzeugen. Im Gegensatz zur Gentechnik, die im Wesentlichen das Ziel hat, ein Gen von einem Organismus in einen anderen zu transferieren, will die synthetische Biologie komplett neue und künstliche biologische Systeme schaffen.
Eine der grundlegenden Techniken, die synthetische Biologen hierzu einsetzen, ist die DNA-Assemblierung, mit der sie zum Beispiel Plasmid-Bibliotheken, Mehr-Komponenten-Systeme oder synthetische Zellen fertigen. Um die hierzu nötigen Gen-Konstrukte herstellen zu können, entwirft man zunächst mithilfe einer Software die entsprechenden Gensequenzen und teilt diese in kleinere überlappende Sequenzen mit etwa 200 bis 1.500 Basenpaare (bp) auf, um sie leichter synthetisieren zu können. Diese sogenannten Synthons werden anschließend zu längeren DNA-Sequenzen assembliert, in einen Expressionsvektor kloniert und schließlich auf Funktionalität getestet. Funktionieren sie nicht wie gewünscht, wiederholt man das Ganze.
Assemblierung mit Restriktionsenzymen...
Für die Assemblierung wurden in den letzten Jahren verschiedene Techniken vorgestellt. Eine der ersten war die GoldenGate-Klonierung, die Sylvestre Marillonets Gruppe am Biozentrum in Halle entwickelte (PLoS ONE 3(11): e3647). Marillonets Methode nutzt Typ-IIS-Restriktionsenzyme (Bbsl, BsmBI, Bsal), die außerhalb der Erkennungssequenz schneiden, sowie T4-DNA-Ligasen für den In-vitro-Zusammenbau multipler DNA-Fragmente. Typ-IIS-Restriktionsenzyme produzieren vier Basenpaare lange, nicht palindromische Überhänge, wodurch die einzelnen Fragmente nahtlos verknüpft werden, ohne „Narben“ zu hinterlassen (seamless cloning). Da in dem ligierten Produkt keine Schnittstellen mehr vorhanden sind, kann man Verdau und Ligation simultan durchführen. Das korrekt ligierte Produkt kann hierdurch nicht mehr von Restriktionsenzymen geschnitten werden, die Reaktion ist also irreversibel.
...oder mithilfe homologer Enden
Kurz nach Marillonet veröffentlichte Daniel Gibson vom Craig-Venter-Institut zusammen mit seinen Kollegen eine Assemblierungs-Technik, die auf der Hybridisierung homologer Enden basiert (Nat. Methods, 6(5):343-5). Wie bei klassischen Homologie-basierten Klonierungs-Methoden, etwa der Overlap-Extension-Klonierung, werden die homologen Enden mithilfe einer PCR an den linearisierten Vektor angehängt. Eine T5-Exonuklease zerkaut anschließend die 5‘-Enden und erzeugt komplementäre, überhängende klebrige Enden (sticky ends), die von einer DNA-Polymerase sowie einer Taq-DNA-Ligase bei 50°C verknüpft werden. Die Gruppe um Gibson synthetisierte mit der Methode das gesamte Genom eines Mycoplasma mycoides-Erregers (JCVI-syn1.0) mit 1,08 Mb.
Ähnlich wie die Gibson-Assemblierung funktionieren auch andere Homologie-basierte Klonierungs-Verfahren wie zum Beispiel Sequence and Ligation-Independent Cloning (SLIC), Circular Polymerase Extension Cloning (CPEC) und Seamless Ligation Cloning Extract (SLiCE).
Ebenfalls von Marillonets Gruppe stammt die Modular Cloning (MoClo)-Technik, eine Weiterentwicklung der Golden-Gate-Klonierung (PLoS One, 6:e16765). Auch dieses Verfahren nutzt Typ-IIS-Restriktionsenzyme, die außerhalb der Erkennungssequenzen schneiden und kompatible Überhänge erzeugen, ohne Rückstände zwischen den DNA-Fragmenten zu hinterlassen. Mit MoClo können bis zu sechs DNA-Fragmente in einem Reaktionsgefäß miteinander verbunden werden. Das System besteht aus drei Klonierungs-Vektoren (Level-0, -1 und -2), die für die jeweiligen Assemblierungs-Schritte genutzt werden. Zu Beginn werden grundlegende DNA-Fragmente (Promoter, untranslatierter Abschnitt, kodierende Sequenzen, Terminatoren, et cetera) der Level-0-Plasmide mit einer PCR amplifiziert und in den Zielvektor kloniert. Bis zu sechs Level-0-Konstrukte können miteinander verknüpft werden und ein Level-1-Konstrukt bilden. Im nächsten Schritt werden sechs Level-1-Elemente zu einem Level-2-Element verschmolzen, wodurch mehrere Gene miteinander verbunden werden können. Theoretisch könnte dies endlos so weiter gehen. In der Realität streikt jedoch der Wirtsorganismus, in der Regel E. coli, ab einer bestimmten Größe des Vektors und lässt sich nicht mehr mit diesem transformieren. Dennoch ermöglicht die Methode den Zusammenbau von Multi-Gen-Konstrukten beziehungsweise Bibliotheken, die in weiteren Schritten, etwa für die industrielle Produktion von Metaboliten, verwendet werden können.
Zu den durch Typ-II-Endonukleasen vermittelten Assemblierungs-Systemen zählt auch die von einer chinesischen Gruppe entwickelte MASTER (Methylation-Assisted Tailorable Ends Rational)-Assemblierungs-Technik (Nucleic Acids Res., 41(8):e93). Mit dieser können mehrere DNA-Sequenzen narbenfrei und Sequenz-unabhängig verbunden werden. Ein Schlüsselelement der Methode ist die Endonuklease MspJI, die Eigenschaften von Typ-IIM- sowie Typ-IIS-Endonukleasen kombiniert. MspJI erkennt Methylierungs-spezifische Sequenzen mit einer Länge von vier Basenpaaren und schneidet die DNA außerhalb dieser Erkennungssequenz. Die MASTER-Technik benötigt aber nach wie vor Restriktionsenzyme und eine PCR-Amplifizierung. Für das Zusammensetzen längerer DNA-Sequenzen ist sie daher ungeeignet.
Auch der von Geoff Baldwins Gruppe am Imperial College London entwickelte sogenannte Biopart Assembly Standard for Idempotent Cloning (BASIC) basiert auf dem Verdau mit Typ-II-Restriktionsendonukleasen (Methods Mol. Biol.,1472:79-91). Auf den Verdau folgt hier die Ligation an Oligonukleotid-Linker mit langen einzelsträngigen Überhängen, welche die spätere Position sowie Ausrichtung innerhalb des endgültigen Konstrukts definieren. BASIC arbeitet äußerst genau: Bei vier Assemblierungen liegt die Genauigkeit bei 100 Prozent bei sieben immerhin noch bei 90 Prozent.
Oligo-Clips
Ohne Verdau funktioniert dagegen die sogenannte Paper Clip-Methode, mit der DNA-Fragmente aus bestehenden Bibliotheken in nahezu jedem Plasmid assembliert werden können (Nucleic Acids Res. 42, 20:e154). Dazu sind lediglich vier spezielle Oligonukleotide (Clips) für jedes DNA-Fragment nötig, die dessen Richtung im fertigen Konstrukt vorgeben. Die Clips werden durch die Ligation von Oligonukleotid-Paaren hergestellt und mithilfe einer PCR oder durch Zellextrakt-vermittelte Rekombination assembliert. Sind die Clips fertig, kann man mit ihnen bis zu sechs DNA-Fragmente in beliebiger Reihenfolge schnell und effizient zusammensetzen.
Um eine noch höhere Standardisierung zu erreichen, entwickelten Reshma Shetty, Drew Endy und Thomas Knight vom Massachusetts Institute of Technology die sogenannten BioBricks (J. Biol. Eng., 2:5). Diese Bio-Backsteine werden am 5‘-Ende von einer Prefix- und am 3‘-Ende von einer Suffix-Sequenz flankiert, die zum Beispiel beim BioBrick-Assemblierungs-Standard „10“ drei, beim sogenannten Freiburger-Standard vier Restriktionsschnittstellen aufweisen. Werden zwei BioBricks verbunden, entsteht ein größerer BioBrick mit den gleichen Prefix- sowie Suffix-Restriktionsschnittstellen an beiden Enden. Die Assemblierung von Gen-Konstrukten wird hierdurch deutlich vereinfacht und beschleunigt.
Ähnliche Standardisierungen wurden auch für Plasmide und Vektoren eingeführt, um diese in verschiedenen Systemen beziehungsweise Organismen ohne große Anpassungen einsetzen zu können. Ein Beispiel hierfür ist die Standard European Vector Architecture (SEVA), bei der grundlegende Bausteine, etwa Selektionsmarker oder Replikationsursprünge (ORI), von standardisierten Restriktionsschnittstellen flankiert werden, um sie einfacher klonieren zu können.
Klonieren mit Null-Background
Bei all diesen Methoden muss man die DNA-Fragmente zuerst auf einem Plasmid-Vektor unterbringen, der anschließend in Bakterien transferiert wird, welche die Plasmide vervielfältigen. Ein Nachteil ist hierbei, dass nicht alle Insertionen von DNA-Fragmenten in die Vektoren perfekt funktionieren und die erfolgreichen Insertionen zeitaufwendig selektiert werden müssen. Eine neue Technik, die Stefan Schusters Gruppe von der Universität Bayreuth entwickelte, kommt jedoch ohne den aufwendigen Selektions-Schritt aus (Sci Rep.;9(1):2980 sowie LJ Online https://www.laborjournal.de/editorials/1718.php „Klonieren mit Zebra“).
Die Methode, die sich Zero-Background Vector oder kurz (ZeBRα) nennt, nutzt einen Vektor mit einem Toxin kodierenden Selbstmord-Gen. Werden die DNA-Fragmente erfolgreich in den Vektor eingebaut, wird das Gen ersetzt. Läuft bei der Insertion aber etwas schief, bleibt das Toxin-Gen intakt und dessen Genprodukt tötet das E. coli-Bakterium. Damit ist garantiert, dass nur E. coli-Bakterien, die das DNA-Fragment enthalten, Kolonien bilden. Das mühsame Kolonien-Picken kann man sich hierdurch sparen.
Synthetisieren statt assemblieren
Trotz Standardisierung und ausgeklügelter Klonierungs-Techniken bleibt die Assemblierung von DNA-Fragmenten zeitaufwendig, und was die entstehenden Konstrukte betrifft, teilweise unvorhersehbar. Zumindest für kleine Konstrukte könnte sie aber bald gar nicht mehr nötig sein: Durch sinkende Preise und eine sehr hohe Genauigkeit wird die DNA-Synthese zu einer immer attraktiveren Alternative. Was bereits möglich ist, zeigt die vollständige Synthese eines funktionierenden synthetischen Hefe-Chromosoms mit fast 300.000 Basenpaaren durch ein Team von der Johns Hopkins University (Science 344(6179):55-8).
Für Konstrukte mit mehr als 2.000 Basenpaaren ist die DNA-Assemblierung mit weiter verfeinerten und standardisierten Verfahren aber das Mittel der Wahl – und wird es vermutlich auch noch einige Zeit bleiben.
Letzte Änderungen: 08.05.2019