Unsichtbare Konformere im Rampenlicht
Festkörper-NMR
Henrik Müller
(08.02.2021) Funktionale Konformere eines Biomoleküls existieren manchmal nur für Mikrosekunden, tragen nur Zehntelprozente zu dessen Konformationsensemble bei und bewegen ihre Atome nicht mal ein Ångström. NMR-Relaxationsdispersion macht sie sichtbar – in Lösung wie im Festkörper.
Spiegeln starre 3D-Modelle von biologischen Makromolekülen die Wirklichkeit wider? Verstaubten Schlüssel-Schloss-Prinzipien zufolge ja. Warum finden sich in der Hälfte aller humanen Proteine dann ungeordnete Regionen, die über simple Schleifen als Scharniere zwischen Sekundärstrukturelementen hinausgehen?
Tatsächlich ist jedes dritte humane Protein zu einem Drittel unstrukturiert. Intrinsisch ungeordnete Proteine verbringen ihr Dasein sogar komplett ungefaltet in einem dynamischen Ensemble interkonvertierender Konformere ohne globales Energieminimum. Eine traditionelle 3D-Architektur erwerben sie nur im enthalpisch oder entropisch favorisierten Wechselspiel mit Interaktionspartnern. Ihre Funktion lässt sich nicht mehr mit einfachen Schlüsseln und starren Schlössern erklären.
Solche heterogenen Strukturen können weder kristallographisch noch kryoelektronenmikroskopisch erfasst werden. Entweder widersetzen sie sich der Kristallisation oder tragen nur zu einem Bruchteil zum Beugungsmuster bei. Als Konsequenz enthalten fünfundsiebzig Prozent der Proteine in der Proteindatenbank (rcsb.org) Bereiche ohne Elektronendichte (J. Biomol. Struct. Dyn. 24(4): 325-42).
Proteindynamik wird sichtbar
Das einzige biophysikalische Verfahren, das sowohl über Struktur als auch Dynamik aufklärt, ist die Kernspinresonanzspektroskopie (NMR). Sie beleuchtet Phänomene auf einer Zeitskala von Pikosekunden bis Minuten: von der Diffusion von Megadalton-Komplexen über allosterische Konformationsänderungen bis zur Vibration chemischer Bindungen. Die Summe aller NMR-Observablen macht in der Theorie die Enthalpie- und Entropiebeiträge jedes Atoms zur Thermodynamik und Kinetik des Gesamtsystems zugänglich.
Rasmus Linser und seine Arbeitsgruppe in der Physikalischen Chemie der Technischen Universität Dortmund entwickeln NMR-Methoden, die über starre Grundzustände hinausblicken: „Die Mitglieder einer Familie verwandter Proteine verfügen über ähnliche enzymatisch aktive Taschen. Ein Familienmitglied kann deshalb oft nicht selektiv über diese adressiert werden, sondern Interaktionspartner müssen allosterisch agieren. Hierfür verwendet die Natur manchmal Minderheitenkonformere, die im Konformationsensemble des Proteins selten vorkommen und spezifisch von Bindungspartnern erkannt werden.“
Minderheitenkonformere tragen nur mit Zehntel- bis Hundertstelprozenten zur Gesamtpopulation bei. Entsprechend herausfordernd ist ihre Vermessung. Ihr direkter Nachweis gelingt auch mit der NMR nicht.
Linser erklärt aber: „Grundzustands- und Minderheitenkonformere tauschen in bioenzymatischen Prozessen oft im Mikro- bis Millisekunden-Bereich hin und her. Jede Konformation hat für jeden Atomkern eine spezifische chemische Umgebung und somit Resonanzfrequenz zur Folge. Wechselt die Konformation, ändert sich auch die Resonanzfrequenz eines betroffenen Atomkerns. Die Summe aller Frequenzen bildet gewichtet nach den Häufigkeiten ihrer Konformation das Gesamt-NMR-Signal.“
Der Einfluss dieses chemischen Austauschs auf das NMR-Signal lässt sich mit bestimmten Pulssequenzen unterdrücken, fährt Linser fort: „Da sich chemische Austauschprozesse zwischen Konformationen verschieden schnell abspielen können, klopfen die einzelnen NMR-Techniken unterschiedliche Zeitskalen ab. Für langsame Prozesse im Bereich weniger Hundert Mikrosekunden bis vieler Hundert Millisekunden eignen sich Refokussierungspulse, während schnellere Prozesse Spinlock-Pulse notwendig machen.“
Auf der Refokussierungsseite ist die Carr-Purcell-Meiboom-Gill(CPMG)-Relaxationsdispersion (RD) die beliebteste Methode der Lösungs-NMR, bestätigt Linser. „Bei ihr werden 180-Grad-Pulse eingestrahlt, um den transversalen Magnetisierungsabfall des NMR-Signals zu refokussieren und die destruktive Interferenz von Austauschprozessen hinauszuzögern. Das gelingt, wenn die Pulse dichter aufeinanderfolgen als der Austauschprozess im Protein vonstattengeht.“
„Die Spinlock-Pulse der R1ρ-Relaxationsdispersion dagegen refokussieren Magnetisierung nicht, sondern halten sie entlang einer Richtung fest“, beschreibt Linser die zweite Art an RD-Pulssequenzen. „Dadurch verzögern auch sie den Magnetisierungsabfall, was ebenfalls umso besser funktioniert, je stärker die Pulse relativ zur Zeitskala des Austauschprozesses sind.“
Infos zu seltenen Konformeren
Warum stecken Lebenszeiten von NMR-Signalen voller Informationen über Minderheitenkonformere? „Weil im Abfall des NMR-Signals nicht nur der Grundzustand von Atomkernen codiert ist“, weiß Linser. „In Abwesenheit von Minderheitenkonformeren ändern sich Relaxationsprofile bei stärkeren oder dichteren Radiofrequenzpulsen nicht. Tun sie es doch, ist mehr als eine Konformation vorhanden. Indem NMRler diese Abhängigkeiten der RD-Profile mit dem Bloch-McConnell-Gleichungssystem quantifizieren, können sie die Austauschraten, die Zeitskala des Konformationswechsels, die Häufigkeiten zugrundeliegender Konformationen und die Unterschiede in ihren chemischen Verschiebungen extrahieren. Daraus lassen sich Strukturänderungen hin zum unsichtbaren Zustand rekonstruieren.“
Wie Relaxationsdispersion unsichtbare Zustände messbar macht, demonstrierte zum Beispiel die Arbeitsgruppe von Marius Clore am National Institute of Health, Maryland, USA. Mit 15N-CPMG-RD charakterisierte sie die Wechseldynamik zwischen den drei Grundzuständen von MinE. Dieses Proteindimer sorgt während der Mitose ausschließlich in der Zellmitte für niedrige Konzentrationen an MinCD-Komplex, sodass dieser nur dort die Maschinerie des mehrere Dutzend Proteine umfassenden Divisoms nicht länger inhibiert und ihm erlaubt, eine Scheidewand aufzubauen. Während eines Teilungszyklus konvertiert MinE dafür von einem sechssträngigen β-Faltblatt zwischen vier α-Helices in eine Konformation mit abgelösten α-Helices in ein viersträngiges β-Faltblatt. Als wären solch tiefgreifenden Umfaltungen zwischen drei Grundzuständen nicht genug, nimmt MinE laut Clores Team vier zusätzliche Minderheiten-Konformationen an. Diese existieren zwischen 500 bis 1.700 Mikrosekunden und sind zwischen 0,2 bis 16 Prozent besetzt. Wie sie von einem Grundzustand zum nächsten führen, erklären die NMRler in einem PNAS-Paper (116 (51): 25446-55)
Vielleicht noch verzwickter ist es, die Dynamik in Festkörpern zu messen, wie zum Beispiel in kristallinen Proteinen, fibrillären Proteinen und Membranproteinen. Warum das kein Widerspruch ist, erklärt Linser: „Selbst im Kristallgitter liegen Proteine weiterhin in wässriger Umgebung vor und sind nur durch wenige Kristallkontakte zu Nachbarmolekülen fixiert. Lösungs-NMR-Relaxation gibt zudem nur Wechselwirkungen wieder, die schneller als die Rotationskorrelationszeit sind mit der Proteine um ihre eigene Achse rotieren. Oder es müssen sich für eine Messung die chemischen Verschiebungen der Atomkerne zwischen Konformation A und B ändern. Im Festkörper spielen dagegen weder Rotationskorrelationszeiten noch chemische Verschiebungen eine Rolle. Tatsächlich ist jegliche Bewegung über Relaxationsphänomene abgreifbar.“
Protonen im Fokus
„Dynamikmessungen im Festkörper wurden in den letzten Jahren revolutioniert“, schwärmt Linser. „Bis 2010 dienten nur 15N- und 13C-Markierungen als Reporter für Proteindynamik. Das dichte Protonennetzwerk von Proteinen aber konnte man nicht nutzen, da es keine aufgelösten Signale lieferte. Dank partieller Deuterierung und ultraschnellem Magic Angle Spinning sind Protonen jetzt endlich im Rampenlicht der Festkörper-NMR.“
Der Austausch von Wasserstoff- durch Deuterium-Atome während der rekombinanten Überexpression eines Proteins verdünnt die Anzahl dipolarer Kopplungen in seinem Protonennetzwerk in den spektroskopisch auswertbaren Bereich. Die chemischen Eigenschaften von Biomolekülen ändern sich dadurch nicht.
Magic Angle Spinning (MAS) indes ist in der Festkörper-NMR seit jeher unentbehrlich, da es deren spektroskopische Arbeitsgrundlage schafft. Was die Brownsche Molekularbewegung für Proben in Lösung, ist MAS für Festkörper. Ohne MAS fände sich in Festkörper-NMR-Spektren nur ein nicht aufgelöstes, unbrauchbares Signal aus der Summe aller anisotrop verbreiterten Resonanzfrequenzen, erklärt Linser: „Rotiert eine Messprobe aber entlang der Raumdiagonale, mittelt das die Anisotropie, also Richtungsabhängigkeit, der chemischen Verschiebung und die meisten dipolaren Kopplungen zwischen Atomkernen heraus. Nach Ausmittelung dieser Wechselwirkungen sind ihre NMR-Kohärenzen langlebiger, was sich in Spektren durch schmale Signale widerspiegelt.“
Einzelne NMR-Signale überlappen also nicht länger, sondern können identifiziert werden. Für eine komplette Ausmittelung muss eine Messprobe zwei- bis dreimal schneller rotieren als die auszumittelnde Interaktion das NMR-Signal beeinflusst. Für Dipolwechselwirkungen direkt benachbarter Protonen müsste die MAS-Rate also mehrere Hundert Kilohertz betragen.
Das ist technisch noch unmöglich. Doch bereits die in den letzten Jahren erreichten 100 bis 130 Kilohertz MAS stellen eine feinmechanische Glanzleistung dar. Für solche MAS-Raten müssen wenige Hundert Nanoliter Probe so in zylindrische Rotorhülsen verbracht werden, dass diese im NMR-Probenkopf gasgelagert schweben und durch seitliches Anblasen ihrer Verschlusskappe auf mehrere Millionen Umdrehungen pro Minute beschleunigt werden können. Infolge von Überschall- und Zentrifugaleffekten benötigen die Rotorhülsen dafür Durchmesser von unter einem Millimeter.
Ultraschnelles MAS macht Protonensignale in kristallinen, fibrillären und Membranproteinen also zuordenbar. Doch löscht es gleichzeitig auch jegliche Information über die Wechselwirkung von Atomkernen aus –und eben diese sind für Struktur- und Dynamikaussagen unentbehrlich. Das Dilemma lösen ausgefuchste Recoupling-Pulssequenzen, die herausgemittelte Wechselwirkungen durch spezifische Pulsabfolgen wiedereinführen. Das mag wie ein Schildbürgerstreich klingen, doch ist es keine neue Strategie der NMR-Gemeinschaft. In den letzten Jahrzehnten entwickelten NMR-Arbeitsgruppen unzählige Wiedereinkopplungsverfahren – oft unter putzigen Abkürzungen wie HORROR, CRAMPS oder DREAM. Jede dieser Pulssequenzen führt nur bestimmte Wechselwirkungen zwischen definierten Atomkernen wieder ein. Resultierende NMR-Spektren enthalten also nur eine spezifische Art an Information.
Linsers Arbeitsgruppe vereint all die genannten Aspekte, indem sie Relaxationsdispersion von Protonen zur Messung von Dynamik im Festkörper anwendet. Ohne Übertreibung katapultiert das die Möglichkeiten der Festkörper-NMR auf ein neues Niveau. Linser ergänzt die Details: „Near Rotary-Resonance Relaxation Dispersion, kurz NERRD, von Protonen stellt nicht nur Information über lokale Fluktuationen zur Verfügung, wie es 15N-Relaxationsexperimente für 1H15N-Kopplungen seit Jahren vermögen: Es sondiert auch regionale Bewegungen innerhalb des Proton-Proton-Netzwerks des gesamten Proteins. Weil dipolare 1H1H-Kopplungen die stärksten Wechselwirkungen im Festkörper sind, können wir so von jedem Proton aus bis zu etwa acht Ångström weit blicken.“ Diese Reichweite zusammen mit der Reichhaltigkeit messbarer Wechselwirkungen erinneri an die Bedeutung des Kern-Overhauser-Effekts (NOE) in der Lösungs-NMR: NOEs sind dort der Schlüssel zu hochaufgelösten Distanzinformationen.
Wie NERRD funktioniert, verdeutlicht Linser so: „Ein Spinlock-Puls mit der gleichen Taktung wie die MAS-Frequenz – also beide beispielsweise 60 Kilohertz – koppelt ausgemittelte anisotrope Interaktionen wieder ein. Sind betroffene Atomkerne starr, verfügen also über diskrete Rotationsfrequenzen, verkürzt sich die Lebensdauer ihrer NMR-Signale extrem, wenn die Resonanzbedingung exakt getroffen wird. Schwingt aber der magnetische Dipol zwischen beweglichen Atomkernen ähnlich schnell hin und her wie die Rotorfrequenz, addieren sich Rotor- und Eigenbewegung. Hierdurch wird die Rotationsbewegung des Kerns kurzzeitig beschleunigt oder verlangsamt. Ein beweglicher Atomkern erfüllt die Resonanzbedingung dann schon bei 50 und 70 Kilohertz. Der Bereich der Rotorresonanz verbreitert sich also je nach der Beweglichkeit des Atomkerns. NERRD-Profile unterscheiden sich entsprechend. Indem wir die RD-Profile mit analytischen Gleichungen für Relaxationsraten modellieren, die die Modulation der Lebensdauer von NMR-Signalen durch MAS und Radiofrequenzpulse berücksichtigen, können wir aus ihnen Ordnungsparameter für jeden Aminosäurerest extrahieren.“
Rätselhafte Ligandenerkennung
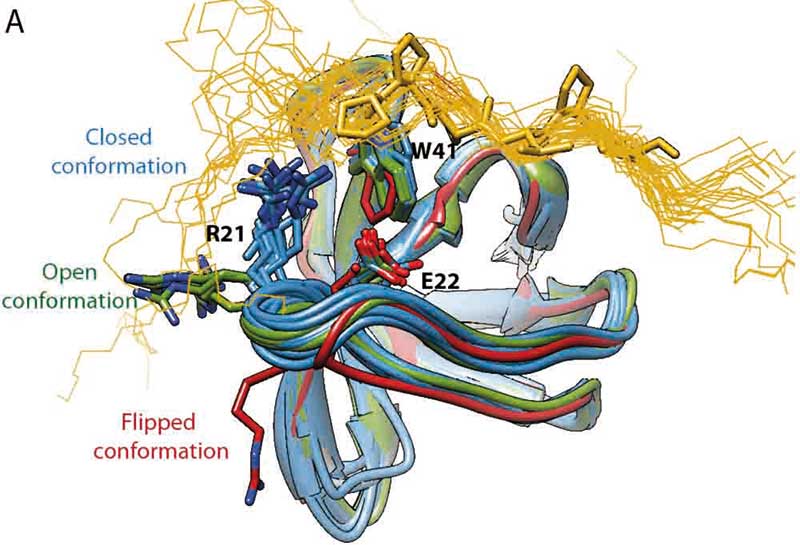
Was sich aus Ordnungsparametern lernen lässt, demonstrierte Linsers Arbeitsgruppe an einem der bestuntersuchten NMR-Modellproteine – der 62 Aminosäurereste langen Alpha-Spektrin SH3-Domäne. Dreihundert humane SH3-Domänen spielen tragende Rollen in Kinasen, Phosphatasen und GTPase-aktivierenden Proteinen in Signalkaskaden des Zellwachstums bis zur Apoptose. Ihre hochkonservierte Tertiärstruktur ist seit Jahrzehnten bekannt. Wie die konformationell heterogenen Kontaktflächen ihrer RT- und N-Src-Schleifen aber Liganden erkennen, ist noch immer unverstanden.
Linsers Arbeitsgruppe begutachtete deshalb die Gesamtheit der Rückgrat- und Seitenkettenfluktuationen in mikrokristallinem SH3 mit ihrem Arsenal mehrerer Relaxationsdispersiontechniken. Als beweglichste Reste stellten sich Gln16 bis Thr24 und Ser36 bis Asn38 heraus, also gerade die RT- und N-Scr-Schleifen der bekannten Erkennungsstelle für Liganden. Die daraus gewonnene Einsicht: Die Erkennungsstelle existiert in einem dynamischen Gleichgewicht zwischen sterisch blockierter Grund-Konformation und bindungskompetenter Minderheiten-Konformation. Letztere nimmt sie alle 170 Mikrosekunden mit einer Wahrscheinlichkeit von 22 Prozent ein (ChemPhysChem. 19 (1): 34-9).
Indem die NMRler experimentelle 15N- und 1H-NERRD-Profile mit simulierten Profilen Hunderter SH3-Kristallstrukturen für potenziell angeregte Zustände verglichen, konnten sie einen Mechanismus der Liganderkennung formulieren. Zuerst bricht die Kation-π-Wechselwirkung zwischen der partiell positiven Seitenkette von Arg21 und der aromatischen Seitenkette von Trp41 auf und erlaubt es Arg21, zu rotieren. Hierdurch flippt die Peptidbindung zwischen Arg21 und Pro20 um 180 Grad in eine geöffnete Konformation. Die Torsionswinkel beider Aminosäurereste gehen von einer konventionellen rechtsgängigen α-Helix in eine ungewöhnliche linksgängige α-Helix über und Arg21 blockiert die Bindungstasche von SH3 jetzt nicht länger. Die Konformationsänderung findet also schon in Abwesenheit von Liganden statt (J. Am. Chem. Soc. 141 (2): 858-69).
Solche strukturdynamischen Modelle stehen erst am Anfang. „Noch können wir die 3D-Konformation angeregter und schwach besetzter Zustände nicht direkt aus Dynamikmessungen rekonstruieren“ so Linser. „In der Lösungs-NMR liefern chemische Verschiebungen in Kombination mit residualen dipolaren Kopplungen bereits Einblicke in die Struktur angeregter Zustände. In der Festkörper-NMR lassen sich chemische Verschiebungen, Austauschraten und Häufigkeiten von Minderheitenkonformeren nur mithilfe komplementärer Daten, etwa aus der Röntgenkristallographie, und mit Molekulardynamik-Simulationen in strukturelle Bilder überführen. Doch wir lernen täglich dazu, wie wir Lösungs- und Festkörper-NMR in einen Gesamtansatz integrieren können, um die strukturellen und mechanistischen Details unsichtbarer Konformere zu rekonstruieren.“
Letzte Änderungen: 08.02.2021