RNA-Interferenz (RNAi)
von Katharina Hien (Laborjournal-Ausgabe 07, 2002)
Wollten sie nicht immer schon eine intensiv pinkfarbene Petunie in ihrem Balkonkasten haben? Nein? Jedenfalls bemühten sich vor etwas mehr als 10 Jahren einige Forscher der Frühlingspflanze eine noch intensivere Pink-Tönung zu verleihen und injizierten ein zusätzliches Pigment-Gen. Die transgenen Petunien blühten jedoch alles andere als pink, nämlich weiß und gefleckt. Das eigene und das eingeschleuste homologe Gen unterdrückten sich gegenseitig - Posttranscriptional Gene Silencing (PTGS) oder Co-Supression war entdeckt.
Zur gleichen Zeit häuften sich ähnliche Erkenntnisse bei Pilzen (Quelling), Würmern, Fliegen und Mäusen. Allen ist gemeinsam, dass eine in die Zelle geschleuste RNA-Sequenz die Konzentration der homologen zelleigenen mRNA stark absenkt, obwohl das betroffene Gen immer noch transkribiert wird. Am Ende erwiesen sich jedoch nicht die fremden einsträngigen sense- oder antisense-Sequenzen für das Gene-Silencing direkt verantwortlich. Vielmehr entdeckten Andrew Fire und Co. 1996 in Caenorhabditis elegans doppelsträngige RNA als den eigentlichen Trigger für die sogenannte RNA Interferenz (RNAi).
 Praktische Methode
Praktische Methode
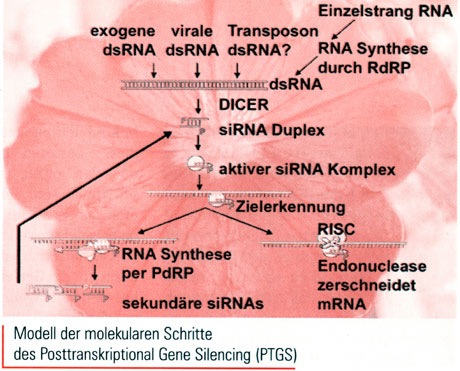
Mittlerweile nutzt man RNAi um bekannte Gene auf unkompliziertere Art auszuschalten als beispielsweise mit Gene-Targeting per homologer Rekombination. Eine in die Zielzelle eingeschleuste dsRNA mit homologer Sequenz zu einer zelleigenen mRNA startet den Vorgang. Dicer, eine ATP-abhängige Ribonuclease, zerschneidet im ersten Schritt die dsRNA in viele kleine Stücke. Die resultierenden Schnittfragmente bestehen auf jeder Seite aus 21-25 Nucleotiden, die am 3'-Ende jeweils 2-3 Nucleotide überstehen, so genannte small interfering RNA (siRNA). Die Fragmente können doppel- oder einsträngig auftreten. Im zweiten Schritt wandert die siRNA in einen weiteren Enzymkomplex, den in Drosophila nachgewiesenen RNA-induced-silencing-complex, kurz RISC. Diese Endoribonuclease findet anhand der siRNA das komplemehtäre Stück der zelleigenen mRNA und zerschneidet sie (Science 296, S. 1270). Der Prozess legt letztlich die Expression des Gens lahm, von der die mRNA stammt: Es wird zwar noch transkribiert, RNAi baut jedoch die mRNA genauso schnell wieder ab.
Das Forscherherz freut sich natürlich: RNAi macht es möglich, mit wenig dsRNA eine immense Menge mRNA zu knacken, im Gegensatz zur herkömmlichen Methode mit antisense RNA, die beispielsweise eine 1000fache RNA-Menge benötigt um ein einzelnes Gen zu blockieren. Der Verstärkunsmechanismus ist noch nicht völlig geklärt, es gibt jedoch mehrere Theorien. Die Dicer-Ribonuclease schneidet jede dsRNA in zirka 10-20 siRNA. Davon wirkt nun jede einzeln im Silencing-Vorgang und erkennt die mRNA an unterschiedlichen Stellen. Eine weitere Möglichkeit ist, dass eine Art katalytischer Prozess jede siRNA mehrfach nutzt. Es gibt jedoch auch Hinweise in C. elegans, dass die siRNA auch als Primer auf die Ziel-mRNA wirken könnte. Eine RNA-Polymerase könnte dadurch sekundäre siRNA produzieren (Science 296, S. 1263).
Komplizierte Säuger
In Säugerzellen ist alles etwas komplizierter, da in die Zelle geschleuste dsRNA unspezifische Auswirkungen haben kann. Eine dsRNA mit mehr als 30 Basenpaaren bewirkt die enzymatische Zerstörung aller mRNAs und den Stopp der Proteinsynthese, vermutlich zum Schutz vor Infektion (Nature 411, S. 428). inzwischen ist es jedoch Routine, kurze siRNAs direkt zu synthetisieren oder zu bestellen. Damit steigt man im Silencing-Vorgang zwar erst später ein, erzielt jedoch den gleichen Effekt.
Aber RNAi entwickelte sich natürlich nicht zur Erleichterung des Forscherlebens. Es ist vielmehr eine Möglichkeit für Zellen, sich effektiv vor Viren zu schützen. Ein Hinweis dafür ist die Tatsache, dass in einem Tiervirus ein Repressor für RNAi gefunden wurde. Am deutlichsten zeigt sich dieser Zusammenhang jedoch in Pflanzen, die sich gegen regelmäßig auftretenden Virenbefall schützen müssen (Science 296, S. 1263).
RNAi ist sogar mit dem Immunsystem der Vertebraten vergleichbar. Beide Systeme erkennen eindringende Parasiten, starten eine spezifische immunantwort und verstärken sie. Der Silencing-Vorgang hat in einigen Organismen wie in C. elegans und Pflanzen auch einen systemischen Effekt. Beispielsweise beginnen Nachbarn einer infizierten Blattzelle ebenfalls die Immunantwort zu starten. Doch wie erkennt die Zelle eine Vireninvasion? Was gibt ihr das Signal, RNAi anzuwerfen?
Ein kleines Immunsystem
Die meisten Viren speichern und replizieren ihr Genom als RNA. Sie können mit drei verschiedenen "lnfektionsgenomen" daherkommen: mit Doppelstrang RNA, messenger-sense RNA (positiver Strang) und antisense RNA (negativer Strang). Sicherlich ist die dsRNA des Virus als solche bereits ein Trigger um RNAi zu starten. Doch wie erkennt die infizierte Zelle fremde Einzel-RNA-Stränge? Paul Ahlquist vermutet, dass eine zelleigene RNA-abhängige RNA-Polymerase (RdRP) das einsträngige Viren-Genom erkennt, in doppelsträngige RNA umwandelt und damit wiederum den RNAi-Vorgang startet (Science 296, S. 1270).
PTGS scheint eine uralte Erfindung der Eukaryoten zu sein, läuft jedoch nicht in allen Organismen völlig identisch ab. Es gibt beispielsweise strukturelle Unterschiede in RdRPs, in Drosophila- und menschlichen Zellen scheinen diese überhaupt keine Rolle zu spielen, und Nematoden benötigen sekundäre siRNA. Aber die für PTGS benötigten Proteine konnten (außer in der Bierhefe) in Protozoen, Pflanzen, Pilzen und Tieren nachgewiesen werden. Archaebakterien und Prokaryoten dagegen fehlt dieser Proteinsatz ganz (Science 296, S. 1265).
Letzte Änderungen: 20.10.2004






