Faszinierende Spezialistin
Tobias Ludwig
(08.05.2020) REGENSBURG: Biochemiker um Christoph Engel ergründen die beachtliche Spezifität der RNA-Polymerase I
und nähern sich der Antwort auf die Frage, warum diese trotz aller Ähnlichkeit zu den anderen Polymerasen doch so anders ist.
Transkription ist ein essenzieller Prozess in allen Lebewesen. Permanent müssen Teile der DNA abgeschrieben werden, damit die Zelle überlebenswichtige Funktionen ausführen kann. In eukaryotischen Zellen übernehmen die RNA-Polymerasen (Pol) I, II und III diesen verantwortungsvollen Job. Doch nicht jede der drei Polymerasen stieß in der Vergangenheit auf Interesse. „Nach meiner Masterarbeit ist mir aufgefallen, dass es nur von einer der drei Polymerasen überhaupt strukturelle Informationen gibt – das war die Polymerase II. Die Pol I ist aber die eigentlich faszinierende“, erinnert sich Christoph Engel, Professor für Strukturelle Biochemie an der Universität Regensburg. Bereits während seiner Doktorarbeit an der Ludwig-Maximilians-Universität München gelang es ihm, die erste Kristallstruktur der Polymerase I zu lösen. Als Postdoc am Max-Planck-Institut für biophysikalische Chemie in Göttingen tauchte Engel tiefer in die Materie ein und beschäftigte sich mit der Regulation und faszinierenden Effizienz von Pol I.
Doch was ist eigentlich das Besondere an dieser Polymerase? Engel: „Pol I ist ein Enzym mit einem ganz speziellen Transkriptionssystem, das sich entwickelt hat, um nur ein einziges Gen beziehungsweise eine Genklasse zu transkribieren. Alle anderen Gene werden mittels differentieller Regulation von Pol II oder III abgeschrieben.“ Pol I produziert ausschließlich die ribosomale Vorläufer-RNA (prä-rRNA), die dann zu einzelnen rRNAs weiter prozessiert wird. Diese Moleküle sind essenzielle Bestandteile von Ribosomen und somit notwendig, damit die Zelle überhaupt Proteine herstellen kann.
Tupfengleiche Transkription
Trotz ihrer unterschiedlichen Aufgaben ist der Aufbau der drei eukaryotischen RNA-Polymerasen recht ähnlich. Sie bestehen alle aus zwei Hauptmodulen und der zentralen DNA-Bindespalte, variieren jedoch in ihrer Zusammensetzung: Pol I besteht aus 14, Pol II aus 12 und die Pol III aus 17 Untereinheiten. Die eigentliche Transkription läuft bei allen drei Polymerasen gleich ab. Zu Beginn muss der abzuschreibende Doppelstrang des DNA-Abschnitts entzweit, also geschmolzen werden. Dieses Aufschmelzen startet am Promoter. An der stromabwärts gelegenen Transcription Start Site (TSS) beginnt das Enzym dann mit der Transkription.
Große Differenzen zeigen sich jedoch vor dem Start der Transkription. „Bei der Pol II, zum Beispiel, gibt es ein bestimmtes Set an Initiationsfaktoren, die da sein müssen. Dazu gehört unter anderem das TATA Box Binding Protein (TBP), der Transcription Factor II B (TFIIB) und weitere Faktoren“, erzählt Engel. „Bei Pol I gibt es zwar Proteine mit strukturellen Ähnlichkeiten, deren Funktion und Anordnung unterscheidet sich aber grundlegend.“ So sei es bisher auch gescheitert, den Initiationskomplex von Pol I (also den Verbund an Polymerase-Untereinheiten und Co-Faktoren) zu Beginn der Transkription auf Basis der Initiationsstadien von Pol II und III zu modellieren. Zudem wird das Abschreiben der DNA durch diese beiden Polymerasen je nach Bedarf komplex reguliert.
Pol I ist da etwas pflegeleichter, wie Engel erläutert: „Bei Pol I ist die Hauptebene der Regulation die Initiation. Die Polymerase muss nur zur richtigen Stelle rekrutiert werden und fängt dann ohne weitere Energiezufuhr an, die DNA zu schmelzen und äußerst effizient abzuschreiben. Es gibt dann nur ‚An‘ oder ‚Aus‘.“ Diese Regulationsebene sei jedoch besonders wichtig, da schnell wachsende Zellen beispielsweise viele Ribosomen benötigten. Würde die Pol I zur falschen Zeit an- oder ausgeschaltet, könne dies gravierende Folgen haben.
Eine Stufe zurück
Damit die Regensburger Pol I näher unter die Lupe nehmen konnten, mussten sie diese zunächst in ausreichenden Mengen herstellen. Kein triviales Unterfangen wie Engel beschreibt: „Die aus 14 Untereinheiten bestehende Polymerase kann man nicht künstlich herstellen, dafür ist sie zu komplex. Wir können sie derzeit nur aus Hefestämmen aufreinigen, in denen wir eine Polymerase-Untereinheit mit einem Affinitätstag versehen haben. Die Zelle setzt die Polymerase dann korrekt zusammen und wir nutzen den Affinitätstag zur Reinigung.“
Nachdem die Forscher die Polymerase sowie Co-Faktoren isoliert und gereinigt hatten, mussten sie den Initiationskomplex in vitro zusammenbauen. In der Vergangenheit hatten sie bereits die Polymerase und den Co-Faktor Rrn3 gemeinsam inkubiert und dann mit dem Core-Factor-Komplex an ein sogenanntes Promoter-Gerüst angebracht. Die Krux dabei: Das Gerüst besteht aus einem künstlich aufgeschmolzenen DNA-Abschnitt sowie einem dazwischen liegenden RNA-Stück – aber mit diesen sogenannten initial transkribierenden Komplexen lässt sich weder das Aufschmelzen der DNA noch der eigentliche Beginn der Transkription studieren, gibt Engel zu bedenken.
p>Daher verfolgten die Biochemiker, allen voran Doktorand und Erstautor der kürzlich veröffentlichten Studie Michael Pilsl, einen neuen Ansatz (
Nat Commun. 11: 1206). Statt die Polymerase auf ein bereits aufgeschmolzenes Gerüst zu setzen, verwendeten die Regensburger ein etwa 160 Basenpaare langes, doppelsträngiges DNA-Fragment, das neben dem Andocken der Polymerase auch die Bindung der Co-Faktoren
Upstream Activation Factor (UAF) und TBP erlaubte. Diese Faktoren helfen dabei, die Polymerase zum korrekten DNA-Abschnitt zu lotsen und binden weiter stromaufwärts. Den Abschnitt direkt nach der TSS begrenzten die Forscher auf 8 Basenpaare, um die Polymerase in einem frühen Übergangszustand der Transkriptionsinitiation zu stabilisieren. Die Gruppe um Engel konnte nun nicht nur die Interaktion der stromaufwärts gelegenen Co-Faktoren mit dem
Core Factor untersuchen, sondern auch, wie genau Pol I die Promoter-DNA aufschmilzt.
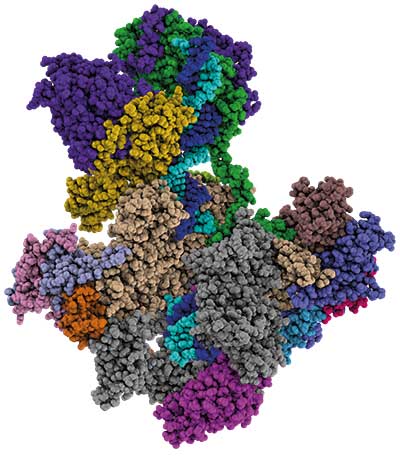
Ein Kryo-Elektronenmikroskop half dem Regensburger Team, den Initiationskomplex auf atomarer Ebene zu betrachten. Dafür säuberten sie die Polymerasekomplexe mittels Größenausschluss-Chromatographie von nicht-assemblierten Bestandteilen und brachten diese dann auf ein sogenanntes Netzchen auf. Das mit einem ultradünnen Kohlenstoffträger versehene Netzchen ermöglichte den Wissenschaftlern, einzelne Moleküle zu separieren.
Die auf dem Träger immobilisierten Proben tauchten die Regensburger in flüssiges Ethan. Dabei werden die Komplexe in glasartigem (vitrifiziertem) Eis eingeschlossen. Die so fixierten Proben untersuchten sie dann unter dem Elektronenmikroskop. Engels Labor schoss über 300.000 Partikelbilder, die das Team in der Folge nach unterschiedlichen Kriterien reduzierte. „Wir haben die Partikel erst zwei- dann dreidimensional klassifiziert und schlussendlich Partikelzustände mit viel Flexibilität aussortiert, da hier möglicherweise beim Einfrieren Untereinheiten verloren gegangen sind“, fasst Engel zusammen. So gelang es den Wissenschaftlern, aus letztlich über 120.000 verbleibenden Partikeln eine Struktur mit einer Auflösung von 3,54 Angström abzuleiten.
Wenn’s wackelt, ist es wichtig
Innerhalb der größtenteils rigiden Struktur des frühen Pol-I-Initiationskomplexes entdeckten Pilsl und Engel dabei immer wieder Abschnitte höherer Flexibilität. „Solche flexiblen Bereiche innerhalb eines Moleküls haben oft biologische Funktionen. Sie können beispielsweise posttranslational modifiziert werden oder interagieren auf eine spezifische Art und Weise“, so Engel. Eine derartige intramolekulare Biegsamkeit fanden die Forscher bei dem Pol-I-Co-Faktor Rrn7. Dieser verfügt über einen Schleifen-Bereich, der wichtig für die korrekte Transkriptionsinitiation ist. Engels Gruppe entfernte die flexible Schleife des Co-Faktors und zeigte, dass die Polymerase das DNA-Muster so nicht mehr effizient abschreiben konnte.
Verwirren und verbiegen
Die Forscher konnten auch erkennen, dass sich die Promoter-DNA, bevor sie von Pol I gebunden werden kann, erst ziemlich verbiegen muss. „Nur wenn die DNA auf diese Weise gebogen werden kann, können alle Kontakte realisiert werden. Es kommt da zu Interaktionen zwischen Polymerase und Core Factor, Core Factor und Promoter-DNA sowie Promoter-DNA und Polymerase“, erklärt Engel.
Zusammenfassend zeigten die Ergebnisse, dass Pol I trotz ihrer strukturellen Ähnlichkeit zu den übrigen Polymerasen einen völlig anderen Mechanismus der Transkriptionsinitiation aufweist. Eine Erkenntnis, die vielleicht bei der Klärung der Frage helfen kann, warum ein so spezifisches Transkriptionssystem für eine einzige RNA notwendig und sinnvoll ist.
Für die Zukunft plant Engels Gruppe daher, die Stabilisierung des Gesamtkomplexes zu optimieren, um den kompletten Rekrutierungs- und Initiationsprozess näher untersuchen zu können. „Regensburg ist da ein sehr schöner Standort, vor allem für mich als Nachwuchsgruppenleiter. Die Interaktion mit den beiden großen Biochemielehrstühlen I und III ist unwahrscheinlich fruchtbar für uns, und wir konnten jetzt auch selbst ein modernes Elektronenmikroskop anschaffen“, schwärmt Engel abschließend.
Letzte Änderungen: 08.05.2020