Neujustierung des molekularen Zollstocks
FRET in orthogonalen Systemen
Heinz Langhals
Biowissenschaftler setzen FRET-Systeme als molekulare Lineale ein, wenn sie Entfernungen zwischen Biomolekülen messen wollen. Aber funktioniert das FRET-Lineal tatsächlich so, wie Forscher bisher dachten? Und stimmt die Skalierung?
Die lichtinduzierte Energieübertragung durch Fluoreszenz-Resonanz-Energie-Transfer (FRET), bei dem ein durch Fluoreszenz angregtes Donor-Chromophor seine Energie strahlungslos an ein Akzeptorchromophor weitergibt, entdeckten Wissenschaftler schon vor mehr als hundert Jahren. Sie taten sich aber lange Zeit schwer, den zugrundeliegenden Mechanismus quantitativ zu beschreiben.Erst die Pionierarbeit des deutschen Physikochemikers Theodor Förster brachte 1946 den Durchbruch. Man bezeichnet dieses physikalische Phänomen, auf dem mittlerweile unzählige Assays, Kits und Untersuchungsmethoden in Biochemie und Molekularbiologie basieren, deshalb meist als Förster-Resonanz-Energie-Transfer.
Förster ging bei seinem Konzept für die Energieübertragung davon aus, dass sich die Lichtquanten abgebenden und aufnehmenden Chromophore, analog zu Radioantennen, wie schwingende elektrische Dipole verhalten. Bringt man zwei schwingende Dipole in Nachbarschaft, dann können sie in Resonanz treten und Energie von einem Dipol auf den anderen übertragen. Das funktioniert ganz ähnlich wie bei einem Musikinstrument das durch die Resonanz eines anderen Instrumentes in Schwingungen versetzt werden kann.
Wichtiges Werkzeug
Försters Theorie von den schwingenden Dipolen verknüpft die Energieübertragung mit molekularen Strukturparametern. Sie ist deshalb sowohl für die Untersuchung photophysikalischer Prozesse, wie etwa des Photosynthese-Reaktionszentrums interessant, als auch als Werkzeug für die Untersuchung biologischer Strukturen und deren Änderungen (siehe dazu
LJ 5, 2011, Seite 70).
Erstaunlicherweise haben sich Biochemiker oder Biologen, die in ihren Laboren tagtäglich mit FRET-basierten Assays und Analysen zu tun haben, aber scheinbar noch nie gefragt, ob die Förster-Theorie, auf der ihre FRET-Messungen beruhen, das FRET-Phänomen auch vollständig beschreibt. Nach unseren jüngsten Ergebnissen ist dies nicht immer der Fall (Heinz Langhals et al.,
J. Am. Chem. Soc. 2010, 132, 16777-82).
Die Förster-Gleichung, mit der man die Energietransferrate, (k
FRET) oder einfacher ausgedrückt, die Stärke des FRETs, berechnet, sieht folgendermaßen aus:
Für Biowissenschaftler sind insbesondere die beiden Strukturparameter R
6DA und κ
2 interessant. Dies nicht nur, weil es Biochemie und Biophysik-Professoren geben soll, die ihre Delinquenten in der Diplom- oder Doktorprüfung schon mal nach diesen zwei physikalischen Größen der Förster-Gleichung fragen. Sie sind vielmehr auch die für biochemische FRET-Anwendungen entscheidenden Faktoren. R
DA ist der Abstand von Donor und Akzeptor-Dipol, geht mit der sechsten Potenz in die Formel ein und steht im Nenner. Es ist deshalb einleuchtend, dass der Energietransfer bei einem zu großen Abstand R
DA nicht mehr funktioniert.
Da die Effizienz des FRET-Prozesses sehr empfindlich vom Abstand der Chromophore abhängt, kann man mit der Förster-Gleichung berechnen, wie nah zwei Molekülteile, etwa zwei an einen DNA-Strang gebundene Strukturen, einander sind. Darüber hinaus lassen sich auch winzige Entfernungsänderungen erfassen. So kann man zum Beispiel die Substratbindung eines Antikörpers, der die beiden Chromophore trägt, über die Veränderung seiner Gestalt und der damit verbundenen kleinen Abstandsänderung bestimmen.
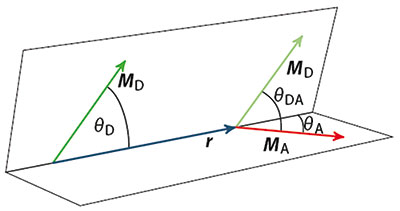
Für unsere Ausgangsfrage ob die Förster-Gleichung den Energietransfer immer richtig wiedergibt, ist jedoch der Faktor κ
2 ausschlaggebend. Er beschreibt die räumliche Orientierung der Dipole zueinander, die das Schaubild oben verdeutlicht, und kann mit der folgenden einfachen Formel berechnet werden:
κ = cos (Θ
DA ) - 3(cos(Θ
A) · cos(Θ
A))
Bei beweglichen Molekülteilen resultiert aus der statistischen Orientierung der Dipole ein Wert für
2 von ungefähr 2/3, den man in der Praxis meist für die FRET-Berechnung verwendet. Nehmen wir aber an, die Dipole stehen senkrecht (orthogonal) zueinander, das heißt die Winkel Θ
DA und Θ
A betragen 90&Deg;, und sie können sich nicht bewegen. Da der Kosinus von 90&Deg; Null ist, sollten nach der Förster-Gleichung auch κ und k
FRET Null sein. Mit anderen Worten: Bei einer orthogonalen Orientierung von Donor- und Akzeptor Dipol zueinander dürfte man nach der Förster-Theorie keinen FRET beobachten.
FRET trotz Verbot
Um dies zu überprüfen, synthetisierten wir ein Donor-Akzeptor Paar das durch ein starres Verbindungsmolekül (Spacer) verbunden ist und dessen elektronische Übergangsmomente orthogonal sind. Zudem steht bei diesem Paar die Verlängerung des Akzeptor-Dipolmoments senkrecht auf dem Mittelpunkt des Donor-Dipols.
Theoretisch sollte κ in diesem Fall den Wert Null annehmen und keine Energieübertragung stattfinden. Bei FRET-Experimenten mit unserem Farbstoff-Paar fanden wir aber genau das Gegenteil: der Donor überträgt die Energie nahezu vollständig auf den senkrecht zum Donor stehenden Akzeptor und benötigt dazu gerade einmal 9,4 Picosekunden. Da wir den zweiten möglichen Mechanismus der Energieübertragung (den so genannten Dexter-Mechanismus) nach zusätzlichen Experimenten ausschließen konnten, bleibt nur der Förster-Resonanz-Energie-Transfer als Erklärung übrig.