Grüne Optogenetik - Optogenetik in Pflanzen
Henrik Müller
(08.04.2021) Lichtsensitive Proteine revolutionierten in den letzten zwanzig Jahren die Erforschung zellulärer
Prozesse in Prokaryoten, Pilzen und vielzelligen Tieren. In Pflanzen verhinderten fehlende Chromophore jedoch ihren Siegeszug. Zumindest bisher.
Welche Eigenschaften sollte die perfekte Untersuchungsmethode haben? Bei Biowissenschaftlern dürften „nicht-invasiv“,‚atoxisch“ und „hochaufgelöst“ ganz oben auf der Wunschliste stehen. Bringen Messverfahren diese Ideale mit, tragen sie das Potenzial in sich, ganze Wissenschaftszweige umzukrempeln. Genau das geschah während der letzten zwanzig Jahren in den Neurowissenschaften. Optogenetische Methoden, mit denen sich molekulargenetisch modifizierte Zellen mit Licht steuern lassen, revolutionierten hier die experimentelle Herangehensweise.
Nature Methods kürte die Optogenetik 2010 zur „Methode des Jahres“. Der Brain Prize veredelte sie 2013 zur „Methode des Jahrzehnts“. Im Herbst 2020 wurde den Optogenetik-Pionieren Peter Hegemann von der Humboldt-Universität zu Berlin, Gero Miesenböck von der University of Oxford und Georg Nagel von der Universität Würzburg der mit 1,2 Millionen US-Dollar dotierte Shaw Prize in Lebenswissenschaften und Medizin verliehen.
Was genau war dem Trio gelungen? Miesenböck sensibilisierte im Jahr 2002 Wirbeltierneuronen für Licht, indem er Rhodopsin aus Drosophila heterolog in Nervenzellkulturen exprimierte. Gleichzeitig entdeckten Hegemann und Nagel das Blaulicht-empfindliche Kanalrhodopsin ChR2 in phototaktischen Grünalgen der Gattung Chlamydomonas (PNAS 100 (24): 13940-45). Dieser Ionenkanal dient den Einzellern als sensorischer Photorezeptor bei schwachem Licht. Im Labor lässt sich mit ihm die elektrische Erregbarkeit von Nervenzellen unmittelbar kontrollieren.
Die Photozyklen von Rhodopsinen unterscheiden sich grundsätzlich. Zwar bestehen alle Rhodopsine aus sieben Transmembran-Helices, die ein Retinyl-Chromophor als protonierte Schiffsche Base kovalent über einen Lysinrest binden. Während eukaryotische Rhodopsine aber Ionenkanäle indirekt über sekundäre Botenstoffe öffnen, bilden Kanalrhodopsine selbst eine Pore durch die Plasmamembran.
Bei den Sehpigmenten vielzelliger Tiere photoisomerisiert 11-cis-Retinal zu all-trans-Retinal. Das hierdurch aktivierte Rhodopsin zerfällt in Chromophor und Opsin, wodurch die Rezeptorzelle über eine mehrschrittige Reaktionskaskade hyperpolarisiert wird. Ohne weitere Belichtung konvertiert eine Retinal-Isomerase den Chromophor zurück in die 11-cis-Konfiguration, welche die Opsin-Komponente zu Rhodopsin regeneriert.
Bindet Retinal dagegen an Kanalrhodopsine, folgt es einem anderen Photozyklus. Georg Nagel, seit 2004 Professor für Molekulare Pflanzenphysiologie und Biophysik an der Universität Würzburg, fasst zusammen: „Mikrobielle Opsine binden kovalent an all-trans-Retinal, das bei Belichtung aber nicht aus 11-cis-Retinal entsteht, sondern zu 13-cis-Retinal isomerisiert. Seine kovalente Bindung zum Opsin bleibt dabei während des gesamten Photozyklus erhalten.“
In der 13-cis-Konfiguration öffnet sich der Ionenkanal, durch dessen 0,6 Nanometer weite Pore mono- und divalente Kationen wie Natrium, Kalium und Calcium ins Zytosol strömen und das Membranpotential innerhalb von Millisekunden ändern. Schließlich relaxiert 13-cis-Retinal zurück in den all-trans-Grundzustand. Das sich hierdurch schließende Kanalrhodopsin unterbricht danach den Ionenfluss.
Warum das für den Experimentator revolutionär ist, brachte Georg Nagel in einem Laborjournal-Essay auf den Punkt (LJ 7-8/2019: Link): „Diese einzigartige Eigenschaft von Kanalrhodopsinen bietet viele neue Möglichkeiten, lebende Zellen oder Organismen durch einzelne Lichtpulse nicht-invasiv zu beeinflussen.“ Schleust man Kanalrhodopsine durch virale Transfektion, Elektroporation oder per Genkanone in Neuronen ein, lässt sich die Aktivität neuronaler Kommunikationsnetzwerke elegant in vitro und in vivo steuern. Auf diese Weise kann man das Verhalten transgener Nematoden, Taufliegen, Zebrafische und Mäuse berührungslos kontrollieren.
Kanalproteine aus Algen
Mittlerweile tragen neben Chlamydomonas auch andere Algengattungen mit Kation-leitenden Kanalproteinen zum optogentischen Werkzeugkasten bei (addgene.org/guides/optogenetics/). Darüber hinaus reagieren synthetische ChR-Chimären, die Transmembran-Helices unterschiedlichen Ursprungs enthalten, nicht mehr nur auf Blaulicht, sondern auf verschiedene Wellenlängen zwischen 450 und 650 Nanometern. Das Gleiche gilt für ChR-Varianten, die durch Punktmutationen modifiziert wurden. Co-exprimiert erlauben es diese, Zellpopulationen gleichzeitig mit Lichtpulsen unterschiedlicher Farben zu kontrollieren. Sogenannte Step-function-Opsine öffnen sich bei einer Wellenlänge und schließen sich bei einer anderen.
Als Gegenspieler für Kationenkanäle fungieren Anion-selektive Kanalproteine, die meist Haloarchaeen entstammen. Mit Grünlicht aktivierbare Chloridkanäle, wie etwa ACR1 der Cryptomonaden-Alge Guillardia theta, regen Neuronen nicht an, sondern inhibieren sie. Wer Neuronen an- oder abschalten will, kann also aus verschiedenen Opsinen auswählen, die einen Großteil des sichtbaren Wellenlängenspektrums abdecken.
Fundamentale Hürden
Während sich tierische Zellen mit optogenetischen Werkzeugen einfach steuern lassen, stehen diesen in Pflanzenzellen fundamentale Hürden im Weg. So besitzen höhere Pflanzen von Natur aus keine Enzyme für die Herstellung von Retinal. Der Chromophor ist aber essentiell für die Photoaktivität von Opsinen. Zwar rekonstituierten Nagels Institutskollegen Dirk Becker und Rainer Hedrich funktionsfähiges ChR2 in Arabidopsis thaliana, indem sie Mesophyllzellen in Medium mit all-trans-Retinal inkubierten (PNAS 117 (34): 20920-25). Weder Gesamtpflanzen noch Sämlinge können aus Kostengründen aber mit den notwendigen 100 µM Chromophor „gefüttert“ werden.
Darüber hinaus lassen sich mikrobielle Rhodopsine nur schwach in Pflanzenzellen exprimieren und finden nur selten ihren Weg in die Plasmamembran. Vor allem benötigen Pflanzen natürlich Licht zur Energiegewinnung – optogenetische Werkzeuge werden hierdurch ungewollt daueraktiviert.
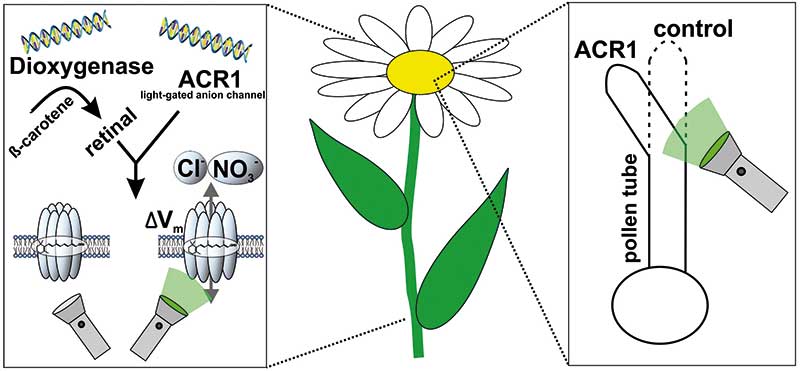
Die Arbeitsgruppe von Georg Nagel ließ sich hiervon jedoch nicht beeindrucken und fand für jedes dieser Probleme eine Lösung (Nat. Plants 7(2): 144-51). Pflanzen synthetisieren zwar kein Retinal, dafür aber dessen Vorstufen. Carotinoide wie α-, β- und γ-Carotin sowie β-Cryptoxanthin dienen ihnen als Lichtsammelpigmente, die Sonnenenergie an die Reaktionszentren der Photosynthese weiterleiten – oder wirken anti-oxidativ, indem sie durch Quenchen vor Sauerstoffradikalen schützen. Vor allem können sie aber in Retinal gespalten werden.
Um diese Spaltung zu erreichen, schleuste Nagels Arbeitsgruppe ein 2009 erstmals beschriebenes Enzym des unkultivierten marinen Bakteriums 66A03 in Pflanzenzellen ein (J. Biol. Chem. 284(23): 15781-93). Diese β-Carotin-15,15‘-Dioxygenase (MbDio) wandelt Carotinoide in Gegenwart von Sauerstoff in all-trans-Retinal um, wenn etwaige Substrate mindestens 35 Kohlenstoffatome und einen unsubstituierten β-Jonon-Ring aufweisen. Alle oben genannten Carotinoide kommen dafür in Frage.
Bereits der erste Versuch, MbDio transient in der Tabakpflanze Nicotiana benthamiana zu exprimieren, gelang. Mehrere Doktoranden um Nagels Postdoc Shiqiang Gao transformierten Agrobacterium tumefaciens mit einem MbDio-Expressionsplasmid, infiltrierten die Bodenbakterien in N.-benthamiana-Blätter und verfolgten die MbDio-Expression mithilfe eines co-transformierten gelb fluoreszierenden Proteins (YFP). Nach der flüssigchromatographischen Aufreinigung des Blattmaterials fand die Arbeitsgruppe Spuren von Retinal in Massenspektrogrammen.
Da Pflanzen Carotinoide in Chloroplasten synthetisieren, nutzten die Pflanzenforscher daraufhin eine 2017 konstruierte Chloroplasten-Transit-Peptidsequenz RC2 (Sci. Rep. 7: 46231), verfrachteten ein RC2-MbDio-YFP-Konstrukt zielgerichtet in die Plastiden und konnten sich über Retinal-Konzentrationen von 140 Nanogramm pro Gramm Blattmaterial freuen.
Kai Konrad, der zweite Seniorautor des Projekts und Arbeitsgruppenleiter am Würzburger Lehrstuhl für Molekulare Pflanzenphysiologie und Biophysik, fasst zusammen: „N.-benthamiana-Blätter sind in zellbiologischen Pflanzenlaboren als transientes heterologes Expressions-System beliebt. Dank simpler Protokolle stellten wir schnell fest, dass sich tatsächlich Retinal in vivo produzieren ließ. Daraufhin wechselten wir zu N. tabacum, einem etablierten System für die stabile Expression von Fremdproteinen.“ Die Würzburger inokulierten Tabaksamen mit Agrobacterium tumefaciens, die ein RC2-MbDio-YFP-Konstrukt beherbergten, kultivierten die heranwachsenden Pflanzen unter Rotlichtbedingungen und fanden in ihren Chloroplasten ähnlich viel Retinal wie bei transienter Expression.
Stabil transformierte Tabakpflanzen wuchsen ebenso gut wie ihr Wildtyp. MbDio beeinflusste weder die Konzentration an Carotinoid-Vorstufen, noch den Gehalt an Chlorophyll a und b. Offensichtlich störte das marine Enzym den Pflanzenmetabolismus nicht. Ermutigt von der erfolgreichen Chromophor-Produktion wandten sich die Pflanzenphysiologen dem eigentlichen optogenetischen Werkzeug zu. Nicotiana hatten sie natürlich mit Bedacht gewählt, erklärt Konrad: „Wir schätzen Tabak nicht nur wegen der Expressions-Protokolle. Seine Wurzelhaare und Pollenschläuche sind auch perfekte Modellsysteme, um den Zusammenhang zwischen Ionenkanal-Aktivität und Pflanzenwachstum zu untersuchen. Besonders die Rolle von Anionenkanälen interessiert uns.“
Monster-Konstrukt
Deshalb erweiterten Nagel et al. ihr RC2-MbDio-YFP-Konstrukt nacheinander um gleich fünf Bestandteile: den mit grünem Licht aktivierbaren Chloridkanal ACR1 aus Guillardia theta (GtACR1), einem selbst-schneidenden Peptid (P2A) des Porzinen Teschovirus 1, zwei Signalpeptiden (E, T) des in Säugern verbreiteten Kaliumkanals Kir2.1 sowie einem chimären Oberflächenmarker humanen und bovinen Ursprungs (LR). Laut Konrad bietet die Monster-Fusion RC2-MbDio-P2A-LR-GtACR1-T-YFP-E große Vorteile gegenüber co-transformierten Plasmiden: „Separate Konstrukte erschweren die Überprüfung der Expression und Membranlokalisation. So brauchten wir N. tabacum nur einmal stabil transformieren, die Transformanden auf ihre YFP-Fluoreszenz screenen und homozygote Pflanzen identifizieren.“
Georg Nagel resümiert: „Dieses Zusammenspiel mehrerer Signalpeptide unterschiedlicher Spezies war nicht zu erwarten, ist aber sehr erfreulich. Oft bin ich erstaunt, wie viel Gemeinsamkeiten bei weit entfernten Organismen zu finden sind!“ So unterbindet P2A die Translation einer Glycin-Prolin-Peptidbindung an seinem eigenen C-Terminus. Entsprechend exprimierten transgene Tabakpflanzen zwei getrennte Proteine: MbDio mit der restlichen P2A-Sequenz als C-Terminus und LR-GtACR1 mit einem N-terminalen Prolin. Keiner der zusätzlichen Aminosäure-Reste störte die zelluläre Prozessierung oder Funktionalität ihrer Proteine.
Die Membranverankerung von GtACR1 erwies sich als größere Herausforderung. Die Kir2.1-Signalpeptide für ER-Export (E) und Membranlokalisation (T) verbesserten zwar die Expression von GtACR1-T-YFP-E-Plasmiden in Xenopus-Oozyten. Transformierten die Würzburger aber Tabakpflanzen, fanden sie keinerlei YFP-Fluoreszenz in deren Epidermiszellen. Die Co-Expression von MbDio und GtACR1 half ein wenig. Vervielfacht wurde die Oberflächenexpression von GtACR1 aber erst durch das chimäre Signalpeptid Lucy-Rho (LR) aus dem N-Terminus des Oberflächenmarkers LRRC32 regulatorischer T-Zellen (Lucy) und dem N-Terminus bovinen Rhodopsins (Rho).
GtACR1 integrierte sich also endlich in die Plasmamembran epidermaler Blattzellen und MbDio synthetisierte Retinal in Chloroplasten. Doch funktionalisierte der Chromophor auch das Kanalrhodopsin? War der Cryptomonaden-Kanal imstande, Zellen höherer Pflanzen bei Belichtung zu depolarisieren?
Ließen die Pflanzen-Biophysiker transgene N.-tabacum-Setzlinge nicht unter Rotlicht, sondern bei Tageslicht wachsen, verringerte sich deren Photosyntheserate und sie verwelkten. Gleichzeitig produzierten sie doppelt so viel epikutikuläres Wachs auf ihren Blättern. Strömten also Anionen durch Tageslicht-aktivierte GtACR1-Kanäle aus Epidermiszellen heraus, weshalb die Tabakpflanzen einen osmotisch getriebenen Wasserverlust durch mehr Oberflächenwachs zu unterbinden suchten?
Elektrophysiologische Messungen legten dies nahe. Im Ruhestand sperren pflanzliche Zellmembranen zehnmal mehr Anionen im Cytoplasma ein als in zwei bis fünf millimolarer Konzentration extrazellulär vorliegt. Entsprechend sind sie um bis zu 200 Millivolt hyperpolarisiert. Belichteten Nagels Mitarbeiter Mesophyllzellen transgener N.-tabacum-Pflanzen fünf Sekunden mit grünem Licht, depolarisierte das deren Membranspannung um bis zu 120 Millivolt.
Tatsächlich erhöhten alle Wellenlängen zwischen 490 und 540 Nanometern das Membranpotential um mindestens 60 Millivolt. Tabakpflanzen, die nur MbDio exprimierten, reagierten dagegen kaum auf gleichartige Lichtpulse. GtACR1 verleibte sich seinen Chromophor also nicht nur ein, sondern quittierte dessen Photoisomerisierung ordnungsgemäß mit Porenöffnung und depolarisierten Membranpotentialen.
Dank der Pionierarbeit von Nagels und Konrads Kooperationsprojekt sind optogenetische Werkzeuge auch der Botanik zugänglich. Sowohl die Chlorophylle in Lichtsammelkomplexen als auch Bilin- und Flavin-abhängige Phytochrome, Cryptochrome und Phototropine pflanzlicher Lichtrezeptoren weisen Absorptionsmaxima im blauen und roten Spektrum auf. Kanalrhodopsine, welche die dazwischenliegende, hundert Nanometer weite grün-gelbe Lücke nutzen, könnten unser pflanzenphysiologisches Verständnis also vervielfachen.
Einen ersten Machbarkeitsnachweis lieferten die Würzburger gleich mit. Kai Konrad erklärt: „Die mit am schnellsten wachsenden Pflanzenzellen sind männliche Gametophyten, sobald sie Pollenschläuche ausbilden. Ändern sich die Richtung oder die Geschwindigkeit ihres Wachstums, können wir das binnen Minuten quantifizieren. Außerdem wissen wir, dass ihr Spitzenwachstum an Ca2+-abhängige Proteinkinasen gebunden ist, die ihrerseits Anionenkanäle kontrollieren. Bisher fehlte uns aber ein experimenteller Beweis, dass aktivierte lokale Anionenkanäle das Spitzenwachstum auch steuern.“
Licht-gesteuerte Pollen
Deshalb beschossen die Würzburger N.-tabacum-Pollenkörner per Genkanone mit MbDio-GtACR1-YFP-Plasmiden. Illuminierten sie transgene Pollenschläuche danach für zehn Minuten mit Grünlicht, hörten sie auf zu wachsen. Schalteten sie das 532-Nanometer-Licht wieder ab, wuchsen die Pollenschläuche weiter. Laut elektrophysiologischer Messungen induzierte ihre Belichtung eine Membran-Depolarisation um fünfzig Millivolt und Photoströme von zwanzig Nanoampere. Offensichtlich kontrollierte GtACR1 tatsächlich die Wachstumsdynamik der Pollenschläuche. Doch steuerte es das Spitzenwachstum auch räumlich?
Belichteten die Pflanzenphysiologen transgene Pollenschläuche asymmetrisch, wuchsen die Pflanzenzellen von der Aktivierungsseite weg. Wildtyp-Zellen ließen sich dagegen nicht von der Belichtung beirren. Damit war der Beweis erbracht: Der Efflux von Anionen und die nachfolgende Änderung des Membranpotentials lenken die Dynamik und Richtung wachsender Pollenschläuche.
Welcher der beiden Effekte ursächlich ist, brachten die bisherigen Experimente allerdings nicht zutage, erklärt Nagel: „Wir würden gerne auch andere Opsine, etwa depolarisierende Kationenkanäle, in Pollenschläuchen exprimieren. Noch ist uns das nicht gelungen.“ Konrad ergänzt: „Der limitierende Faktor sind aber nicht technische Hürden, sondern optogenetische Werkzeuge, die wir gerade erst für die Pflanzenforschung erschließen. Die Sequenzierung ganzer Genome in allen Domänen des Lebens identifiziert immer neue putative Opsine, die kloniert und elektrophysiologisch charakterisiert werden wollen. In Zukunft wird eine Palette lichtgesteuerter Ionenkanäle und Photoschalter unterschiedlicher Anregungsspektren, Leitfähigkeiten und Licht-Sensitivitäten zur Verfügung stehen. Das ist vergleichbar mit GFP, nach dessen Entdeckung eine Vielzahl ähnlicher Fluoreszenzproteine gefunden oder erzeugt wurde.“
Synthetische Biologie und Botanik rücken weiter zusammen.