Dem Goldstandard auf den Fersen - Isothermale Amplifikations-Verfahren
Mario Rembold
(10.11.2021) Beim Nachweis von Nukleinsäuren galt die PCR lange Zeit als Maß aller Dinge. Doch isothermale Amplifikations-Methoden können es inzwischen mit dem Klassiker aufnehmen.
Die PCR gilt als Goldstandard zum Nachweis von DNA oder zuvor revers transkribierter RNA. Allerdings ist die altbewährte Methode auch umständlich, denn man benötigt einen Thermocycler, der die Temperatur innerhalb eines PCR-Zyklus sehr schnell anpasst. Die Geräte sind teuer und oft auch unhandlich, außerdem dauert das Abarbeiten aller Zyklen schon mal ein oder zwei Stunden – je nach Länge des nachzuweisenden Targets muss man der Polymerase ausreichend Zeit für die Amplifikation der DNA lassen. Erst danach können die Doppelstränge wieder aufgeschmolzen werden.
Isothermale-Amplifikations-Verfahren laufen dagegen bei gleichbleibender Temperatur ab – im Idealfall müssen im Verlauf der Amplifikation auch keine Puffer gewechselt werden. Schon 1990 entwickelten Thomas R. Gingeras und Kollegen am Salk Institute Biotechnology in San Diego mit der Self-sustained Sequence Replication (3SR) eine Amplifikationstechnik, die konstant bei 37 Grad Celsius abläuft – die Forscher wiesen damit HI-Viren nach (Ann. Biol. Clin. 48(7): 498-501). Ein Primer, der um eine Promotor-Sequenz verlängert ist, an die sich die T7-RNA-Polymerase anlagern kann, bindet bei 3SR zunächst an das RNA-Target. Nach Zugabe von Reverser Transkriptase entsteht DNA. Anschließend wird die RNA verdaut, und ein zweiter, ebenfalls mit einer T7-PromotorSequenz versehener Primer bindet in Gegenrichtung an die DNA. Danach transkribiert man beide Stränge mithilfe der Promotor-Sequenz sowie durch Zugabe von T7-RNA-Polymerase wieder zu RNA. Es entstehen zwei RNA-Stränge, die wiederum revers transkribiert werden. Jedes Mal verdoppelt sich dabei die Menge an DNA in der Probe.
Obwohl kein Temperaturwechsel notwendig ist, muss man zwischen den einzelnen Schritten RNA verdauen, um RNA-DNA-Duplets aufzulösen. Anschließend wird die RNA wieder synthetisiert, sodass man ebenfalls Zyklen abarbeitet.
Auch zirkuläre Templates lassen sich isothermal vermehren: Zufallsprimer binden am einzelsträngigen Ring, eine stangverdrängende Polymerase verlängert sie. Die Reaktion läuft wieder und wieder um den Template-Ring herum und erzeugt eine einzelsträngige und immer länger werdende lineare DNA. Auf dem Einzelstrang wiederholt sich etliche Male die Sequenz des Templates. Man bezeichnet diese Technik als Rolling-Circle-Amplifikation (RCA), die Menge synthetisierter DNA wächst jedoch nicht exponentiell, sondern nur linear.
Liegt ein doppelsträngiges Template als Ausgangsmaterial vor, muss der Strang auseinander geschoben werden, während die Polymerase einen Primer verlängert. Man verwendet deshalb strangversetzende beziehungsweise Strand-Displacement-Polymerasen. Bereits 1992 stellten Walker und Kollegen in PNAS eine Strand Displacement Amplification (SDA) vor (89(1): 392-6). Ein doppelsträngiges Target-Template wird bei dieser initial aufgeschmolzen. Anschließend bindet ein Primer, der um eine sogenannte Nicking-Sequenz verlängert ist. Am neu entstandenen Doppelstrang kann hier später ein Nicking-Enzym schneiden. Dieses Enzym durchtrennt nur einen Strang, sodass wieder ein Primer binden kann. Die strangversetzende Polymerase fügt dann komplementäre Basen an und schiebt den anderen Doppelstrang beiseite. Diesem Strang fehlt die Nicking-Sequenz – da der Primer später wieder bindet, wird sie in der nächsten Kopie erneut eingefügt.
Einen Überblick über die Geschichte der isothermalen Amplifikation mit übersichtlichen Schemata liefert ein kürzlich vorab online erschienenes Review von Jörn Glökler et al. (Crit Rev. Biochem. Mol. Biol. 56(6): 543-86).
Die Loop-Mediated Isothermal Amplification (LAMP) ist derzeit die bekannteste Methode zur isothermalen Amplifikation – 60 Prozent der bis 2019 veröffentlichten Publikationen zur isothermalen Amplifikation drehten sich um LAMP, stellen Lisa Becherer und ihre Kollegen vom Institut für Mikrosystemtechnik – IMTEK der Universität Freiburg in ihrem 2020 erschienenen Review fest (Anal. Methods 12: 717-746).
Mitautor an dieser Übersicht zur LAMP ist auch der Biochemiker Sieghard Frischmann, der die Produktentwicklung bei der Mast Diagnostica GmbH leitet, der deutschen Niederlassung der Mast Group Ltd. Schwerpunkt seiner Arbeit ist die Infektionsdiagnostik, für den Erregernachweis optimiert sein Team auch die LAMP.
Konstante Temperatur
LAMP findet bei einer gleichbleibenden Temperatur zwischen 60 und 65 Grad Celsius statt, 63 Grad seien recht typisch, so Frischmann. „Die DNA ist bei dieser Temperatur in einem semistabilen Zustand. Es liegen sowohl doppelsträngige als auch einzelsträngige Abschnitte vor.“ Unter diesen Bedingungen ist einerseits Primer-Annealing möglich, andererseits lässt sich die doppelsträngige DNA auch leicht aufschmelzen.
Als DNA-Polymerase verwendete man in der ursprünglichen LAMP Bst-DNA-Polymerase aus Geobacillus stearothermophilus, die auch in anderen isothermalen Amplifikations-Verfahren eingesetzt wird. Heute nutzt man die Bsm-DNA-Polymerase aus Bacillus smithii. Beides sind Strand-Displacement-Polymerasen.
„Weil die Temperaturen nicht so hoch sind wie bei einer PCR, können wir zeitgleich auch eine Reverse Transkriptase zugeben, um Sequenzen aus RNA-Erregern oder anderen RNA-Targets zu amplifizieren“, fährt Frischmann fort. Dafür muss man keine Puffer wechseln oder pipettieren, bestätigt er. „Das läuft alles in einem Reaktionsansatz ab.“
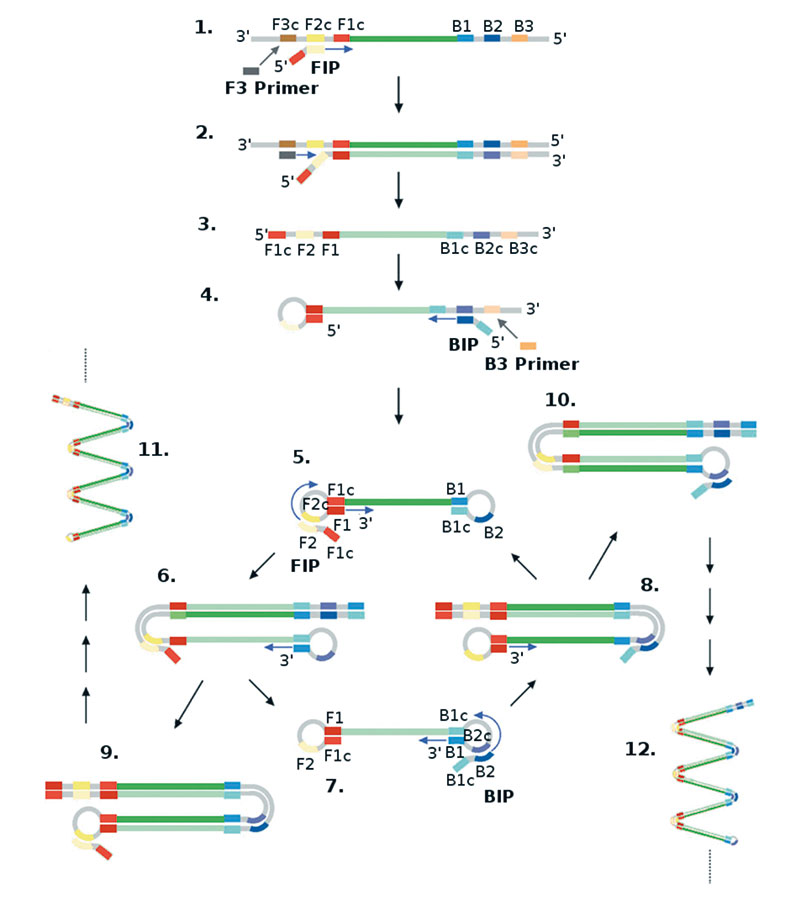
Die Amplifikation geht wie bei der PCR immer von DNA aus. Für die LAMP benötigt man mindestens vier Primer, die Reaktion lässt sich aber durch weitere Primer beschleunigen. In der einfachsten LAMP-Variante setzt man zwei innere und zwei äußere Primer ein, jeweils einen für die Forward- und einen für die Reverse-Richtung. Die inneren Primer sind jeweils um eine kurze Sequenz verlängert – hierzu gleich mehr. Innere und äußere Primer binden an das Template. Weil gleichzeitig auch der benachbarte äußere Primer durch die Polymerase verlängert wird, drängt die Polymerase aufgrund ihrer strangversetzenden Eigenschaft den vom inneren Primer initiierten Einzelstrang beiseite. An diesen Einzelstrang setzen innere und äußere Primer der entgegengesetzten Richtung an, und wieder wird der vom inneren Primer gestartete Strang verdrängt.
Die kurzen Verlängerungen der beiden inneren Primer sind komplementär zu einem Stück des Targets. Der entstandene Einzelstrang paart daher mit sich selbst und formt eine Schleife. Weil auf der anderen Seite ebenfalls ein komplementärer Überhang zu einem anderen Stück Target-Sequenz sitzt, entsteht auch hier eine Schleife. Diese beiden Loops geben dem Strang das Aussehen einer Hantel.
Hantel als Vorlage
Die Hantel ist der Ausgangspunkt für eine exponentielle Amplifikation. Beide inneren Primer binden wieder gleichzeitig, und in der Folge entstehen weitere und auch längere Einzelstränge mit Schleifen, die wieder Vorlage für neue Amplifikationen sind. „In den Loops kann man weitere Primer einsetzen und die Reaktion damit beschleunigen“, so Frischmann. Denn bei der LAMP gibt es keine einzelnen Zyklen, die Reaktionen laufen kontinuierlich und zeitgleich ab. Wo Primer passende Ziele finden, binden sie auch und sorgen für eine Amplifikation. Daher beschleunigt sich die Reaktion, je mehr Primer ins Spiel kommen, häufig sind es sechs bis acht.
Es gibt verschiedene Möglichkeiten, die neu entstehende DNA nachzuweisen. „Die ursprünglich entwickelte LAMP ließ sich visuell auswerten“, weiß Frischmann. „Die Reaktionspuffer waren so konzipiert, dass eine Trübung einsetzte, je länger die Reaktion lief.“ Somit gab es ein zuverlässiges Ja-oder-Nein-Resultat. Man konnte den Grad der Trübung, der mit der Ausgangskonzentration des Targets korreliert, aber auch turbidimetrisch bestimmen. Dies sei aber ein indirekter Nachweis gewesen, denn die Trübung war nicht unmittelbar gekoppelt an die Replikation der DNA.
Hierin sieht Frischmann auch einen der Gründe, warum die klassische PCR die Nase weiterhin vorn behielt, obwohl LAMP bereits im Jahr 2000 von Notomi et al. vorgestellt worden war (Nucleic Acids Res. 28(12): E63). Mit der Real-Time-PCR lässt sich nämlich direkt verfolgen, welche Menge an PCR-Produkt neu entsteht. „In der Molekularbiologie hat man den Anspruch, dass das erhaltene Signal auch unmittelbar widerspiegelt, was sich auf Amplifikations-Ebene gerade tut“, so Frischmann.
Erste Schritte seien dann Farbstoffe gewesen, die in die DNA interkalieren, zum Beispiel SYBR Green. Inzwischen ist diese direkte Kopplung des Signals an die LAMP-Reaktion kein Problem mehr. Vielmehr sind LAMP-Tüftler aus aller Welt derzeit sehr kreativ und entwickeln neue, in der Regel Fluoreszenz-basierte Methoden zur Quantifizierung.
Eine Technik stellte Becherer in ihrer Doktorarbeit vor. Sie benutzt sogenannte Mediator-Displacement-Sonden (MD-Sonden) und spricht daher von der MD-LAMP. Eine Sonde besteht aus einem Forward-Loop-Primer mit einer zum Target komplementären Sequenz sowie einer 5‘-Verlängerung, an die wiederum der Mediator komplementär gebunden ist. Der Mediator wird einzelsträngig abgetrennt, sobald der Primer sein Ziel findet und bindet.
Universeller Reporter
Das eigentliche Signal während der MD-LAMP geht von einem universellen Reporter aus: einem haarnadelförmigen DNA-Stück, an das ein Fluorophor gebunden ist. Allerdings sitzt nahe dem Fluorophor ein Quencher, der die Fluoreszenz unterdrückt. Das 3‘-Ende dieser Haarnadel hängt aber ein wenig über und ist hier komplementär zum freischwimmenden Mediator. Bindet Letzterer an den Reporter, so drängt er den Quencher beiseite und das Fluorophor leuchtet (falls es angeregt wird). Reporter und Mediator sind also unabhängig vom Target. Lediglich der Loop-Forward-Primer, der gemeinsam mit dem Mediator die MD-Sonde bildet, muss an das Target angepasst werden. Das Prinzip der MD-LAMP ist im Review von Becherer et al. erläutert und schematisch dargestellt.
Vergangenes Jahr haben Becherer und Co., ebenfalls in Kooperation mit Frischmann und Mast Diagnostica, einen von der MD-LAMP abgeleiteten Multiplex-Nachweis für zwei bakterielle Krankheitserreger vorgestellt (Emerg. Infect. Dis. 26(2): 282-8). Detektieren wollten die Forscher Treponema pallidum oder Haemophilus ducreyi, beides Erreger der Frambösie, einer Tropenkrankheit, die mit Hautveränderungen einhergeht.
Die Messung mehrerer Targets im selben Ansatz sei lange eine Herausforderung für die LAMP gewesen, erinnert sich Frischmann. „Wir konnten zwar schon 2008 mehrere Primer in eine Probe geben und dreißig bis 40 Subtypen eines Erregers nachweisen, sie ließen sich aber nicht voneinander unterscheiden.“ Hierfür braucht es leicht anpassbare Reporter-Systeme mit unterschiedlichen spezifischen Sonden, die auch individuelle Farbsignale liefern. „Da war uns die PCR immer zwei bis drei Jahre voraus.“
Auch Kontrollreaktionen waren daher nicht im selben Reaktionsgefäß möglich. „Bei einem negativen Ergebnis möchte man ja sicher sein, dass in der Probe keine Substanz enthalten ist, die die Amplifikation inhibiert“, erläutert Frischmann. Heute sei das Problem gelöst, sodass man zu der Probe auch ein Kontroll-Target geben kann, das anzeigt, ob die LAMP stattfand oder nicht. „Inzwischen lassen sich ohne weiteres drei bis vier verschiedene Erreger in einem Tube nachweisen.“
Im Sortiment von Mast Diagnostica gibt es zum Beispiel fertige LAMP-Kits für Salmonellen, Legionellen oder Listerien. Doch der Ökologe im Freiland möchte vielleicht ein ganz ausgefallenes Bakterium schnell in Bodenproben nachweisen, für das es überhaupt keine Fertig-Kits zu kaufen gibt. Stellt sich die Frage, ob LAMP auch im Eigenbau möglich ist.
„Manche bekommen das hin, andere nicht“, konstatiert Frischmann. „Wir bieten aber auch allgemeine generische AmplifikationsKits an. Dann muss man zwar eigene Primer designen, sich aber nicht um Farb- und Pufferlösungen kümmern.“
Gut durchdachte Primer
Da man mit mindestens vier, besser sechs Primern arbeitet, ist die LAMP hochspezifisch. Andererseits besteht immer die Gefahr, dass sich Primerdimere bilden und dann doch ein Signal aufleuchtet, obwohl kein Target in der Probe ist. „Aber für das Primer-Design gibt es wirklich gute Softwarepakete“, beruhigt Frischmann, „dennoch ist natürlich große Sorgfalt geboten, damit die Primer sowohl sensitiv als auch spezifisch sind.“ Will heißen: Die Methode soll so empfindlich sein, dass sie wenig Nukleinsäure-Moleküle erkennt, aber eben nur diese Moleküle und keine andere DNA oder RNA.
„Die designten Primer sollte man immer erst mit negativem Probenmaterial kontrollieren“, mahnt Frischmann. Erst wenn die Resultate zuverlässig negativ sind, könne man sich an positive Proben herantasten und Versuche mit Referenzmaterial durchführen, bei denen die Konzentration der Targets bekannt ist. Nur ein gewissenhaft validierter Test bringe auch zuverlässige Resultate, was aber ohnehin für alle Nachweismethoden gilt.
Wer die LAMP zum Quantifizieren nutzt, sollte sich über einige Besonderheiten im Klaren sein, um die Ergebnisse auch korrekt interpretieren zu können. Frischmann: „Wenn viel Ausgangsmaterial vorliegt, ist das ähnlich wie bei der PCR: Hohe Konzentrationen liefern ein frühes Signal.“
Frischmann erklärt, dass man auch in der LAMP von einem Ct-Wert spricht, wohlwissend, dass hier natürlich keine diskreten Zyklen ablaufen, sondern die Amplifikation in einem kontinuierlichen Prozess geschieht. „Meistens messen wir einmal pro Minute und verwenden diese Zeitschritte als Ct-Wert“, erläutert Frischmann. „Wenn tausend oder mehr Erreger in einer Probe vorhanden sind, taucht das Signal sehr früh auf. Je nach Assay sprechen acht bis zehn Minuten für ein stark positives Ergebnis.“
Zufällig zu schnell
Für mittlere Erregerkonzentrationen nennt Frischmann 18 bis 20 Minuten als Richtwert. Will man jedoch geringe Target-Mengen quantifizieren, ist die LAMP mit Vorsicht zu genießen. Denn die exponentielle Amplifikation beginnt erst, nachdem die erste Hantel generiert wurde. „Es kommt also darauf an, wie rasch diese Startstruktur auftaucht“, so Frischmann. „Entsteht sie aus zufälligen Gründen sehr schnell, kann die LAMP ein stärker positives Ergebnis vortäuschen.“ Sind im Ausgangsmaterial nur zehn Erreger, so dauere es typischerweise zwischen 35 und 40 Minuten, bis ein Signal hochkommt. „Per Zufall kann das aber auch mal in weniger als 30 Minuten passieren.“
An dieser Stelle erwähnt Frischmann die digitale Amplifikation, die sich auch für die LAMP eignet. Mit ihr ist man weit weniger vom Zufall abhängig. Man gibt aufbereitetes Probenmaterial in Form winziger, Öl-umhüllter Tröpfchen auf die Oberfläche eines Chips oder einer Membran. „Man geht davon aus, dass sich in einem Tropfen maximal ein Erreger befindet“, veranschaulicht Frischmann die Idee. Tatsächlich erreicht man diese Voraussetzung erst durch die geeignete Verdünnung der Probe, die umso größer sein muss, je mehr Erreger enthalten sind. In jedem Tröpfchen läuft dann eine LAMP ab. „Das können zwischen 5.000 und 30.000 Tröpfchen sein“, so Frischmann, „und am Ende der Reaktion beobachtet man das Ergebnis mit einer Optik und wertet aus, wie viele Tröpfchen fluoreszieren und eine positive Reaktion anzeigen“.
Digitale LAMP
Die LAMP sei optimal für die digitale Amplifikation, schwärmt Frischmann. „Man kann das natürlich auch mit einer PCR machen, doch unter den hohen Temperaturen leiden die Tröpfchen häufig.“
Zum Nachweis des Humanen T-lymphotropen Virus 1 (HTLV-1) hat ein großes, von Felix von Stetten vom IMTEK geführtes Forscherteam dieses Jahr ein vergleichsweise handliches Komplettsystem vorgestellt, das die Amplifikation in Tröpfchen nutzt. In das Instrument werden Mikrofluidik-Scheibchen eingesetzt, an denen das Gerät die Tröpfchen generiert und die LAMP ablaufen lässt. So kann man das Verfahren auch vor Ort für die Point-of-Care-Diagnostik einsetzen, ohne Proben in große Labore für die Analytik verschicken zu müssen.
HTLV-1 ist ein Retrovirus, das menschliche T-Lymphozyten infiziert und zu einer Leukämie führen kann. In den Tröpfchen des portablen Apparats findet also auch eine Reverse Transkription der viralen RNA statt. Auch hieran hat Frischmann mitgearbeitet, das spezifische Signal entsteht per MD-Sonde – Becherer ist auch hier als Erstautorin gelistet (Micromachines 12(2): 159).
Allgemein gilt die LAMP als sehr robust gegenüber Substanzen im Probenmaterial, die eine klassische PCR stören würden. „Weil die LAMP so unempfindlich auf Inhibitoren reagiert, kann man direkt aus tierischen Kotproben Erreger nachweisen, auch Vollblut lässt sich direkt für die LAMP einsetzen, etwa um Malaria-Erreger zu detektieren“, erläutert Frischmann.
Natürlich wird die LAMP auch im Kampf gegen die Corona-Pandemie genutzt und angepasst. Ein offensichtlicher Vorteil ist die höhere Geschwindigkeit der Amplifikation. Darüber hinaus ermöglicht sie, auch große Mengen Probenmaterial gleichzeitig zu verarbeiten und später zuzuordnen.
Kerstin Ludwig und ihre Kollegen von der Universität Bonn sowie dem Massachusetts Institute of Technology stellen in einem vorab online erschienenen Artikel in Nature Biotechnology ein Verfahren vor, das die LAMP als Vorbereitung für die spätere Sequenzierung nutzt (doi: 10.1038/s41587-021-00966-9). Bei dieser LAMP-Seq werden die Kopien des Targets mit einem eindeutigen Barcode in Form einer DNA-Basenfolge versehen. Für die spätere Sequenzierung kann man die Proben etlicher Spender dann gepoolt analysieren und über den Barcode in der Sequenz später wieder auf die einzelne Person rückschließen.
LAMP und Co. beschleunigen und vereinfachen die Molekularbiologie. Darüber hinaus eröffnen sie auch die Chance, molekulare Diagnostik in Gegenden zu ermöglichen, in denen die medizinische Versorgung weniger gut ist. Und natürlich ist die isothermale Amplifikation auch spannend für jeden Freilandforscher, der nicht überall ein komplettes Labor zur Verfügung hat.