Schalterprotein im Rampenlicht
Larissa Tetsch
(13.05.2022) VILLIGEN (SCHWEIZ): In den extrem lichtempfindlichen Stäbchen der Wirbeltiernetzhaut wandelt ein Ionenkanal Lichtsignale in elektrische Signale um. Seine neu aufgeklärte Struktur könnte helfen, Wirkstoffe zu finden, die defekte Kanalvarianten verschließen können.
Das Wirbeltierauge gehört zu den kompliziertesten Organen überhaupt. Dass wir damit Bilder sehen können, verdanken wir zwei lichtempfindlichen Sinneszelltypen in der Netzhaut: den Stäbchen und Zapfen. Während Erstere für das Sehen im Dämmerlicht verantwortlich sind, dienen die kegelförmigen Zapfen der Wahrnehmung von Farben. Wirbeltiere besitzen zwischen ein und vier Zapfentypen, die jeweils für unterschiedliche Spektralbereiche spezifisch sind. Die Stäbchen sind dagegen besonders lichtempfindlich und können sogar nur ein einzelnes Photon wahrnehmen. Daran beteiligt ist ein Ionenkanal aus der Familie der Cyclic-Nucleotide-Gated(CNG)-Kanäle, der als elektrischer Schalter das Lichtsignal in eine Änderung des Zellmembranpotenzials umwandelt.
Für derartige Schalterproteine interessiert sich Jacopo Marino vom Paul-Scherrer-Institut im schweizerischen Villigen. „Die CNG-Kanäle gehören zu einer Großgruppe von spannungsabhängigen Kanälen“, erklärt der Strukturbiologe. „Das Besondere an ihnen ist, dass sie am cytoplasmatischen C-Terminus eine Bindestelle für 3‘-5‘-zyklische Nukleosidmonophosphate besitzen.“ Im Dunkeln ist die Konzentration des zyklischen Nukleosidmonophosphats GMP (cGMP) in den Sehsinneszellen hoch, die CNG-Kanäle binden cGMP und stehen offen. Ein Lichtsignal aktiviert Rhodopsin, woraufhin cGMP durch die Aktivität der Phosphodiesterase abgebaut wird. Ohne gebundenes cGMP schließen sich die Ionenkanäle. Dadurch verändert sich das Membranpotenzial; es entsteht ein Aktionspotenzial und damit ein elektrisches Signal, das wir letztlich als Lichtblitz sehen.
Neuer Blick auf alten Bekannten
Die tetrameren CNG-Kanäle kommen in verschiedenen Sinneszellen vor. Marino, der seine Doktorarbeit an der Universität Zürich gemacht hat und anschließend als Postdoktorand am Genzentrum der Ludwig-Maximilians-Universität in München beschäftigt war, konzentriert sich mit seiner Forschung auf den Stäbchen-Kanal CNGA1/CNGB1. Dieser besteht aus drei A1-Untereinheiten, die über eine Leucin-Zipper-Region am C-Terminus miteinander verbunden sind, sowie einer strukturell ähnlichen B1-Untereinheit. Letztere bringt ein paar Besonderheiten mit, etwa eine lange, ungeordnete und Glutaminsäure-reiche Region mit noch unbekannter Funktion sowie zwei Bindestellen für das Calciumbindeprotein Calmodulin. Dessen Bindung setzt die Affinität von CNGB1 für cGMP herab und erschwert damit die Öffnung des Kanals.
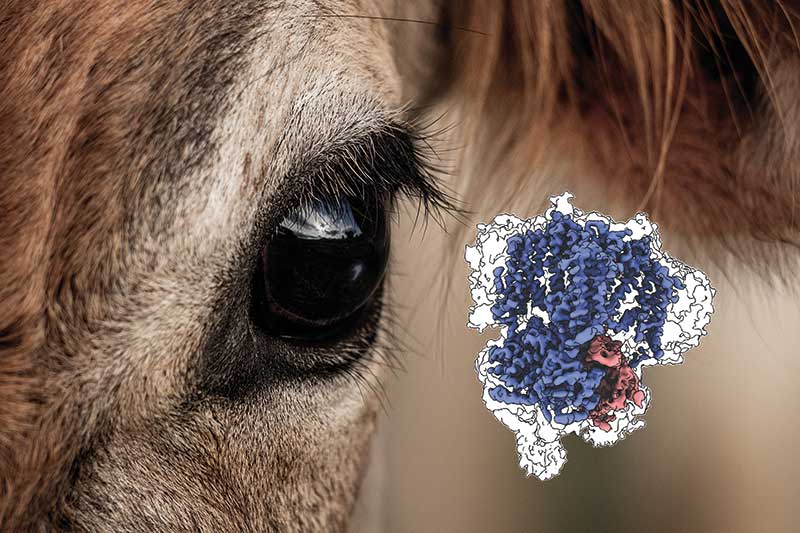
Biochemisch und elektrophysiologisch ist der schon seit rund 40 Jahren bekannte Kanal sehr gut erforscht. Was bisher aber noch fehlte, war die Struktur. Diese ist auch medizinisch von Bedeutung, weil Defekte im Kanal schwere Augenkrankheiten verursachen können, die noch nicht pharmakologisch behandelt werden können. Als Spezialist für die Strukturaufklärung von Membranproteinen wollte Marino diese Wissenslücke schließen. Gemeinsam mit seiner Doktorandin Diane Barret hat er CNGA1/CNGB1 deshalb über Cryo-Elektronenmikroskopie untersucht (Nat. Struct. Mol. Biol. 29(1): 32-9). „Unsere Ergebnisse bestätigen einerseits, was wir aus biochemischen Studien bereits zu wissen glaubten, und liefern andererseits aber auch überraschende Ergebnisse“, freut sich der Strukturbiologe. „Sie bieten jetzt die Grundlage für eine neue Ära der Erforschung der Kanäle.“
Fleißarbeit im Vorfeld
Die größte Hürde für die Strukturaufklärung war, genug Material für die Cryo-Elektronenmikroskopie zu sammeln. Große Proteinmengen gewänne man am einfachsten durch eine heterologe Expression, erklärt Barret. „Bei einem heteromeren Proteinkomplex wie CNGA1/CNGB1 ist das aber schwierig. Man müsste zwei Proteine heterolog exprimieren und diese wieder im richtigen stöchiometrischen Verhältnis zusammenbringen. Das ist zwar machbar, fügt dem Ganzen aber eine weitere Komplexitätsebene hinzu.“
Stattdessen reinigte Barret den Kanal direkt aus der Netzhaut von Rinderaugen – was ziemlich mühsam war. „Der Kanal ist dann schon richtig zusammengebaut, aber man bekommt natürlich nur sehr wenig Material“, so die Erstautorin der Studie. Ein weiterer Nachteil war, dass an die Kanalproteine kein Tag für die Reinigung angebracht werden konnte. Doch auch dafür fand das Forschungsteam eine Lösung, wie Barret berichtet. „Wir haben ausgenutzt, dass die B1-Untereinheit Calmodulin bindet. Damit konnten wir den Kanal aus der Probe fischen.“
Am Ende reichten 80 Mikrogramm reines Kanalprotein aus, um die Struktur mit einer Auflösung von 3,4 Ångström darzustellen. Diese bestätigte, dass der Kanal aus drei A1- und einer B1-Untereinheit aufgebaut ist und dass die drei A1-Untereinheiten am C-Terminus interagieren. Während CNGA1 fast vollständig darstellbar war, fehlten bei der B1-Untereinheit ein paar Regionen vor allem am N-Terminus. „Wir haben die unstrukturierten Abschnitte des Proteins nicht abbilden können, denn diese sind in der Elektronenmikroskopie unsichtbar“, bedauert Barret. Dennoch lieferte die Aufnahme erstmals detaillierte Einblicke in die Struktur eines heteromeren CNG-Kanals und förderte verschiedene Unterschiede zu den strukturell besser erforschten homomeren CNGA1-Kanälen zutage.
Besonders interessant waren die Details der Pore, durch die die Ionen diffundieren. Gleich am Eingang liegt der Selektivitätsfilter, der aus einem Segment der Transmembrandomäne und mehreren Schleifen gebildet wird. Drei Glutamatreste, die von den drei A1-Untereinheiten beigesteuert werden, sorgen mit ihrer negativen Ladung dafür, dass nur positive Ionen wie Calcium, Natrium und Kalium passieren können. „Auf Seiten der B1-Untereinheit liegt anstelle des Glutamats jedoch ein Glycin“, wirft Marino ein. „Dieses ist ungeladen und sehr klein. Deshalb vermuten wir, dass der Durchtritt von Kationen durch die B1-Untereinheit wohl erleichtert wird.“ Unterhalb des Selektivitätsfilters liegt eine Art Tor, das die Pore durch die hydrophoben Aminosäuren Phenylalanin und Valin verschließt. Öffnet sich der Kanal nach der Bindung von cGMP, verschieben sich diese Aminosäuren durch eine große Konformationsänderung, sodass Kationen durchtreten können.
Die größte Überraschung für die Wissenschaftler war aber, dass unterhalb der hydrophoben Engstelle noch ein weiteres Tor liegt. Hier ragt die positiv geladene Seitenkette eines einzigen Argininrests der Untereinheit B1 in die Pore hinein. „Vermutlich behindert sie den Durchtritt der Ionen nicht nur sterisch, sondern auch durch elektrische Abstoßung“, vermutet Marino und fährt fort: „Die Bedeutung des Arginins ist bisher völlig übersehen worden. Die Aminosäure ist zwar auf Proteinebene konserviert, aber ohne Struktur kann man halt nie sagen, wohin die Seitenkette genau zeigt.“ Ob das Arginin komplett weggedreht wird, wenn sich die Pore öffnet, möchten Marino und Barret gerne zukünftig untersuchen.
Variation im Zapfen
Während das Arginin bei CNGB1-Untereinheiten anderer Arten vollständig konserviert ist, fanden die Membranprotein-Forscher bei der CNGB3-Untereinheit des analogen Kanals in Zapfen-Sinneszellen eine interessante Abweichung. Zwar besitzen Arten mit drei oder vier Zapfentypen – Trichromaten wie Primaten und Tetrachromaten wie die meisten Vögel, Fische, Reptilien und einige Säugetiere – in CNGB3 genau wie in CNGB1 ein Arginin an der entsprechenden Stelle. Bei mono- und dichromaten Arten wie Robben und Wale befindet sich an der entsprechenden Stelle jedoch ein Glutamin. „Darüber welchen Unterschied das macht, lässt sich bisher nur spekulieren“, sagt Marino. „Wahrscheinlich wird der Durchtritt aber für die Ionen einfacher, weil die Seitenkette kürzer ist und die Ladung wegfällt.“ Weitere Untersuchungen könnten Klarheit bringen.
Aber wie so oft gilt auch hier: Keine Regel ohne Ausnahme! Bei dichromaten Pferden und Nashörnern findet sich trotzdem ein Arginin, beim vermutlich tetrachromaten neuseeländischen Okarito-Kiwi ein Glutamin.
In ihrem Artikel diskutieren Marino und Barret in diesem Zusammenhang eine interessante Hypothese: Ein codiertes Glutamin lässt sich durch eine Adenosin-Deaminierung auf RNA-Ebene in ein Arginin umwandeln und dieser RNA-Editierungsmechanismus wird in manchen Ionenkanälen im Säugetier-Gehirn tatsächlich genutzt, um die entsprechenden Kanäle weniger durchlässig für Calciumionen zu machen. Könnte so ein Mechanismus auch bei CNGA1/CNGB1 eine Rolle spielen? „Wir finden die Idee ansprechend“, gibt Marino zu, „aber Beweise haben wir noch nicht. Wir müssten dafür die mRNA in der Retina untersuchen und schauen, ob eine bestimmte Base als Mix vorliegt. Das steht auf jeden Fall auf unserer Agenda.“
Ansonsten wollen Marino und Barret als Nächstes unbedingt die Struktur des offenen Kanals analysieren, um beispielsweise herauszufinden, was beim Öffnen mit dem Arginin in der Pore geschieht. Zwar wurde kürzlich bereits die Struktur des offenen CNGA1/CNGB1-Kanals veröffentlicht, aber aufgrund unterschiedlicher Präparationsbedingungen ist der direkte Vergleich schwierig.
Am Ende bleibt zu hoffen, dass die Struktur tatsächlich hilft, Wirkstoffe gegen Augenkrankheiten wie die Retinitis pigmentosa zu finden. „Das ist das ultimative Ziel der Strukturbiologie“, sagt Marino. „Allerdings müssen wir dazu vermutlich industrielle Partner finden, denn die Wirkstoffsuche ist teuer und sehr zeitaufwendig.“